Таймер тим 01 принцип работы: Как проверить таймер оттайки тим 01 – таймер, электронный таймер, ТИМ-01, ТИМ-01Н, принцип работы, электросхема, подключение, проверка, дефекты, ремонт, холодильник, Тольятти, ремонт холодильника, Тольятти, Омск, Томск, Самара, Ульяновск, Новосибирск, Воронеж, Липецк, Барнаул, Нижневартовск, Тюмень, Екатеринбург, Владивосток, Хабаровск, Калуга, Брест, Минск, Киев, Тобольск, Сургут, Ханты-Мансийск, Салехард, Красноярск, Сочи, Новороссийск, Чита, Архангельск, Курган, Уфа, Казань, Москва, Астрахань
Таймер электронный ТЭО-02 (ТИМ-01) режим оттайки 12 часов (в кор 100шт)
Если Вы еще не работаете с нами, оставьте здесь свои данные и мы обязательно свяжемся с Вами
Округ *Центральный федеральный округСибирский федеральный округДальневосточный федеральный округУральский федеральный округПриволжский федеральный округЮжный федеральный округСеверо-Западный федеральный округ
Регион *Белгородская областьБрянская областьВладимирская областьВоронежская областьг. МоскваИвановская областьКалужская областьКостромская областьКурская областьЛипецкая областьМосковская областьОрловская областьРязанская областьСмоленская областьТамбовская областьТверская областьТульская областьЯрославская областьРегион *Агинский Бурятский автономный округАлтайский крайИркутская областьКемеровская областьКрасноярский крайНовосибирская областьОмская областьРеспублика АлтайРеспублика БурятияРеспублика ТываРеспублика ХакасияТаймырский автономный округТомская областьУсть-Ордынский автономный округЧитинская областьЭвенкийский автономный округРегион *Амурская областьЕврейская автономная областьКамчатская областьКорякский автономный округМагаданская областьПриморский крайРеспублика Саха (Якутия)Сахалинская областьХабаровский крайЧукотский автономный округРегион *Курганская областьСвердловская областьТюменская областьХанты-Мансийский автономный округЧелябинская областьЯмало-Ненецкий автономный округРегион *Кировская областьКоми-Пермяцкий автономный округНижегородская областьОренбургская областьПензенская областьПермская областьРеспублика БашкортостанРеспублика Марий ЭлРеспублика МордовияРеспублика ТатарстанСамарская областьСаратовская областьУдмуртская РеспубликаУльяновская областьЧувашская РеспубликаРегион *Астраханская областьВолгоградская областьКабардино-Балкарская РеспубликаКарачаево-Черкесская РеспубликаКраснодарский крайРеспублика АдыгеяРеспублика ДагестанРеспублика ИнгушетияРеспублика КалмыкияРеспублика Северная Осетия-АланияРостовская областьСтавропольский крайЧеченская РеспубликаРегион *Архангельская областьВологодская областьг.
Тип покупателя *Розничный покупательЧастный мастер по ремонтуОптовый клиент
Сообщение
Таймер электронный оттайки ТИМ-01 для холодильников Indesit, Ariston
Артикул C00298587
Производитель: [Россия]
Таймер оттайки для холодильников INDESIT, ARISTON, STINOL и других
Таймер электронный, ТИМ-01Н ВБ А-класс
Коды замен: С00277450 W16002554500, 851161, 277063, 298587, 851086, ТЭУ-01-01, ТЭУ-01-02
Версии таймеров ТИМ-01:
- ТИМ – 01.
 11
11 - ТИМ – 01С
- ТИМ – 01Н
- ТИМ – 01Ф -2
- ТИМ – 01Ф -3
- ТИМ – 01Ф -8
- ТИМ – 01Т -2
- ТИМ – 01Т -3
- ТИМ – 01Т -8
- ТИМ – 01.11 -1
- ТИМ – 01С -1
- ТИМ – 01Н -1
 11
11C 2003г. таймеры ТИМ-01 поставляются со временем паузы 2 мин. (допуск срабатывания отсутствует) вместо 7 мин.
В 2013г. завод холодильников “Индезит” г.Липецк (холодильники Индезит, Аристон) начал устанавливать вместо таймеров ТИМ-01 производства “Протон-Импульс” г.Орел, таймеры производства ФРИДЖЕС ПАРТС г.Липецк модели ТЭУ-01-2 “Унибоб-Л”).
ПРИНЦИП РАБОТЫ ТАЙМЕРА:
Цикл оттайки испарителя начинается через 8 или 12 часов работы компрессора (зависит от класса). При этом время стоянки не учитывается.
Условия начала разморозки и включения нагревателя:
- В морозильной камере температура должна быть ниже – 10 град. С.
- При достижении температуры на термореле в МК +10 град. С цикл оттайки прекращается.
- При первом включении холодильника в сеть и при достижении температуры – 10 град. С в МК на термореле, таймер введет холодильник в оттайку

Помните, что таймер при повторном включении будет помнить о том, что он уже был включен в сеть. Т.е. перевод из режима в режим будет осуществляться при «ВКЛ»/ «ВЫКЛ» положении переключателя и нажатии на кнопку на таймере. «Забудет» он, т.е. счетчики сбросят, не менее, чем через 8 часов
Для таймеров УНИБОБ ручной перевод из режима в режим не предусмотрен. «Память» будет сохраняться не менее, чем 4 часа.
Цена:
669
pуб.
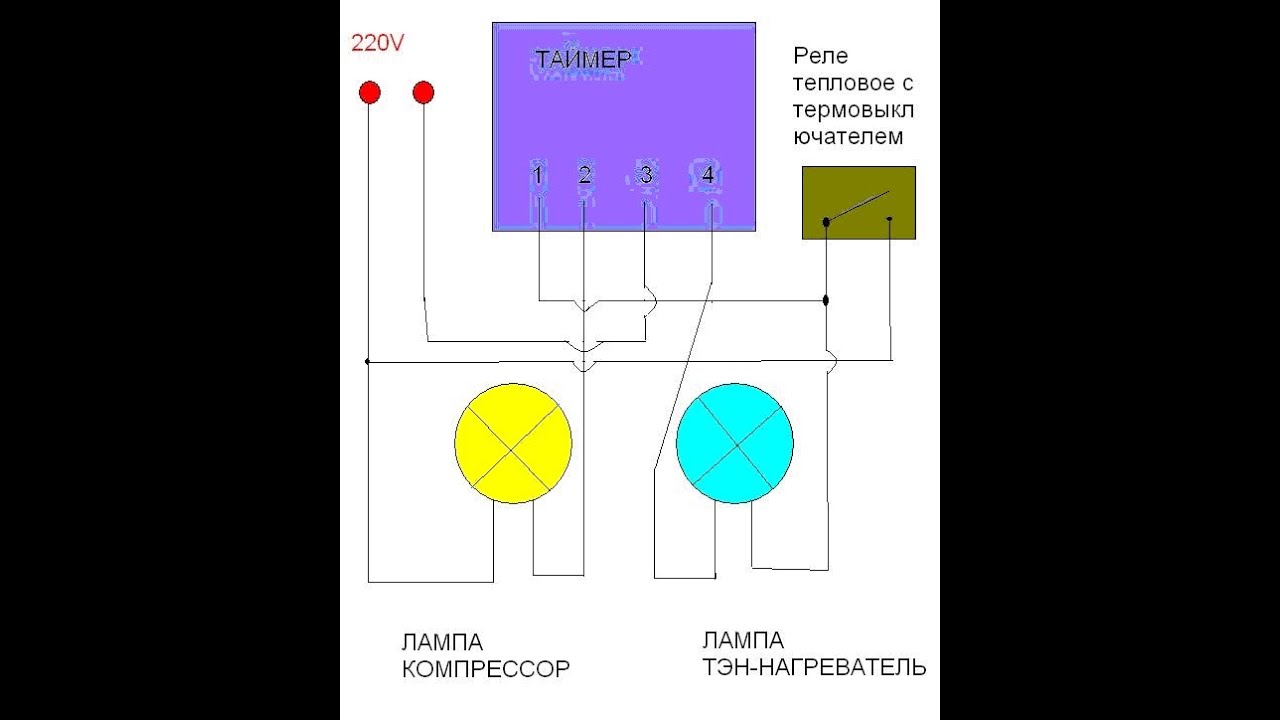
Схема подключения таймера тим 01
ремонт NoFrost таймера холодильников Stinol, Indesit, Ariston
Электронные таймеры оттайки можно условно разделить на 2 большие группы —
ТИМ-01 на микросхеме с маркировкой “ХМ3” и ТЭУ-01 на “Аttiny13”
Понятно, что замена перегоревших резисторов или залипшей кнопки не тема для статьи.
Хочу поделиться способом оживления именно микросхем таймера.
И если неисправные таймеры на “тиньке” со сгоревшим контроллером, как правило, отправляются сразу в мусор,
то часто таймер оттайки типа ТИМ 01 можно вернуть к полноценной жизни.
Если плата вашего неисправного таймера выглядит вот так (фото с Монитора):
И все детали исправны, а сигнал на включение оттайки с выхода 3 микросхемы ХМ3 не поступает,
скорее всего, в обрыве коллекторный резистор.
Внешний резистор номиналом от 4.7кОм до 5. 1 кОм с 8-й ножки на 3-ю излечивает дефект микросхемы.
1 кОм с 8-й ножки на 3-ю излечивает дефект микросхемы.
По-видимому, она КМОП логики и не рассчитана на подключенный к её выходу 3 резистор 1 кОм.
Более 10 шт восстановленных таймеров исправно трудятся уже 2 года.
основная задача устранить проблемы в работе вашего холодильника качественно, в кратчайший срок и по приемлемой цене
- Главная
- электронный таймер ТИМ-01
электронный таймер ТИМ-01
3. После этого восстанавливают целостность схемы холодильника. При необходимости заменяют дефектный таймер
Индезит NBA18FNF Индезит C132NFQ.016
ТИМ-01С
ТЭУ-01-2 – оттайка электронагревателем, сохранение информации о режиме работы при отключении электропитания в течении 8 часов, продолжительность режима активной оттайки определяется тепловым реле, но не более.
РВ-1 электронный таймер предназначен для автоматического управления режимом оттайки
Автор: ides · Опубликовано 14.07.2016 · Обновлено 14.07.2016
Электронный таймер ТИМ- 01 (применяется для управления оттайкой испарителя холодильников No Frost, Full No Frost, Total No Frost — Индезит, Аристон
Оттайка начинается при условии – температура в морозильной камере не выше – 10 С
Окончание оттайки – при температуре на термореле +10 С.
 Время оттайки зависит от количества инея на испарителе.
Время оттайки зависит от количества инея на испарителе. Электронный таймер ТИМ- 01Н
При первом включении холодильника, при достижении температуры в морозильной камере -10 С, таймер входит в режим оттайка – используется для контроля работы системы оттайки.
Оттайка начинается при условии – температура в морозильной камере не выше – 10 С
Окончание оттайки – при температуре на термореле +10С. Время оттайки зависит от количества инея на испарителе
Параметры для проверки:
— ручная установка режима оттайки
— время паузы
— включение холодильного режима
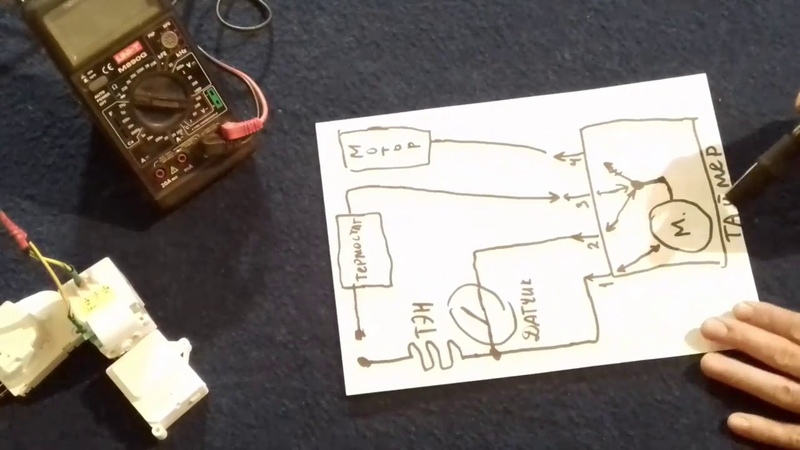
Проверяют в следующей последовательности
2. Отсоединяют провод термопредохранителя от коммутационной колодки, тем самым имитируется размыкание контактов теплового реле, после этого если используется таймер ТИМ -01 – компрессор включается через 7 (+- 3) мин.

3. После этого восстанавливают целостность схемы холодильника. При необходимости заменяют дефектный таймер
Индезит NBA18FNF Индезит C132NFQ.016
ТИМ-01С
ТЭУ-01-2 – оттайка электронагревателем, сохранение информации о режиме работы при отключении электропитания в течении 8 часов, продолжительность режима активной оттайки определяется тепловым реле, но не более….
РВ-1 электронный таймер предназначен для автоматического управления режимом оттайки
Дефекты холодильника Full No Frost
Дефекты перечислены по степени вероятности возникновения. 1 Дефект теплового реле испарителя, в результате чего не подается напряжение на тэн, испаритель полностью затягивает инеем в течении 2, 3 дней после возникновения дефекта
Дефекты системы NO FROST
Дефект представлен на одном холодильнике — сначала произошло произошло отслоение тэна каплепадения, тэн прожег теплоизоляцию, перемерз канал слива конденсата,накопившимся
Принцип работы холодильника Full No Frost
Основное назначение системы – предотвращение образования инея внутри холодильной и морозильной камер за счет принудительного прогона воздуха.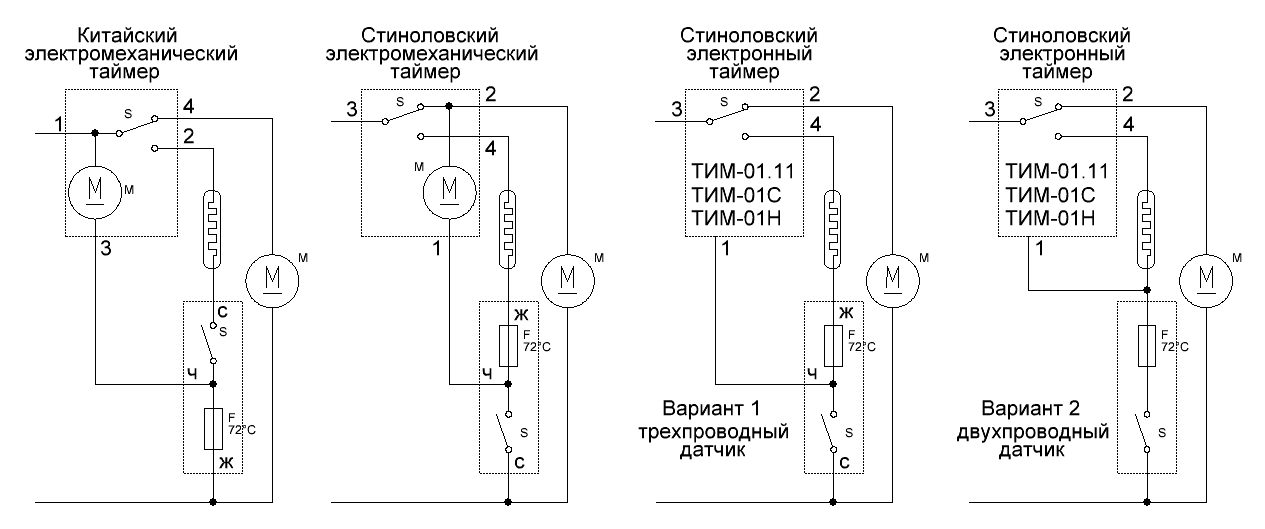
Cроки хранения продуктов при близкриоскопических температурах
Ряд бытовых холодильников оснащен камерами для хранения продуктов в близкриоскопическом состоянии, т.е. при стабильно близких к нулю температурах. Стабильность температуры в процессе хранения значительно влияет на продолжительность хранения, причем в разной степени на различные продукты. Все пищевые продукты можно условно
Для чего нужен таймер оттайки системы NF в холодильнике?
Светлана Викторовна
Очень сильно начал гудеть холодильник, а морозильная камера вообще перестала морозить. Мастер оказался настоящим профессионалом своего дела. За что вам огромное спасибо! Сразу определил причину поломки и быстро ее устранил. Теперь холодильник работает как часы.
24.05.2021Виктор
Сдавал к ним холодильник в ремонт винный холодильник в начале мая, почему-то перестал морозить. Оперативно забрали, за несколько дней починили и вернули назад уже рабочий. По деньгам обошлось недорого, рекомендую!
Оперативно забрали, за несколько дней починили и вернули назад уже рабочий. По деньгам обошлось недорого, рекомендую!
Дмитрий
На заявку отреагировали быстро, специалист позвонил перед приездом. Диагностику и собственно ремонт произвёл качественно и относительно не долго. Техника теперь работает отлично, хочу выразить благодарность за качественные услуги и ответственное отношение.
23.04.2021Виктория Сергеевна
Хочу поблагодарить за хорошую работу мастера Александра! Управился всего за 40 минут, вдобавок ещё и помог настроить систему охлаждения! Приятный парень, на диспетчерской линии тоже замечательные люди. Спасибо что вы есть!
07.03.2021Сергей
Сервис хороший. Перестала работать морозилка. Вызвал мастера. Мастер приехал в течение часа и всё сделал, так что сейчас морозилка и весь холодильник в полном порядке. Спасибо!
14.02.2021Алексей Владимирович
Хочу оставить свой отзыв о работе сервиса. Ребята все приличные и адекватные, решили проблему с моим холодильником во время НГ праздников.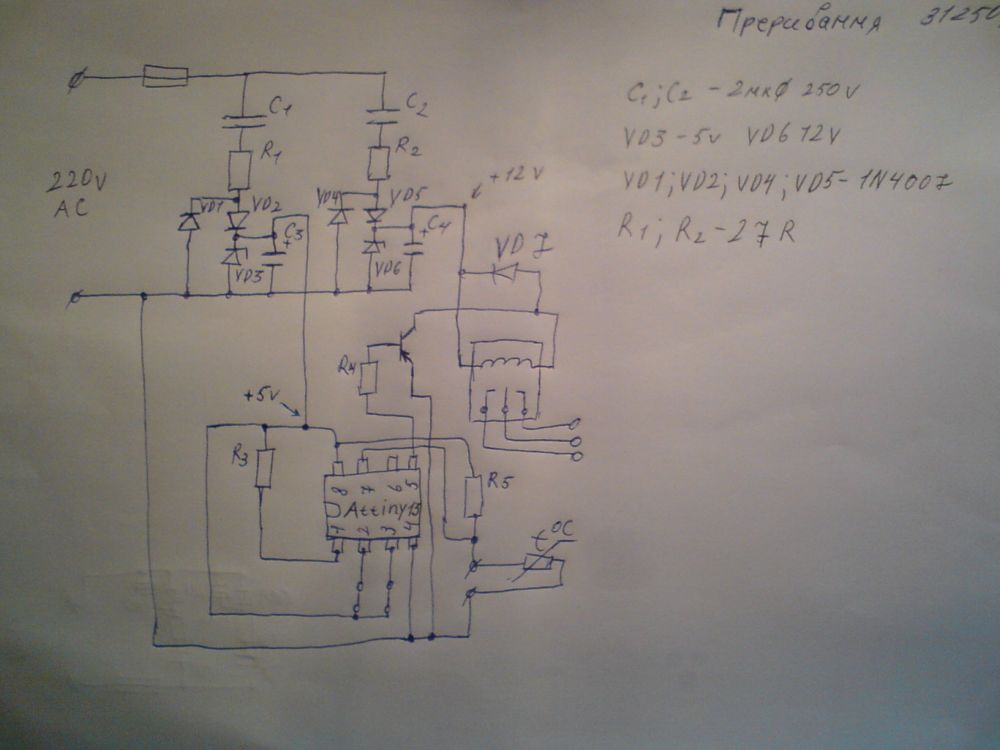 Цена ремонта адекватная. Спасибо.
Цена ремонта адекватная. Спасибо.
Михаил К.
Обращались в эту компанию 31 декабря, на удивление мастер приехал 1 января и к тому же в приличном виде после новогодней ночи))). Рекомендую!
Михаил К.
05.01.2021Юрий
Выручила нас ваш сервис, спасибо! Холодильник много лет работал исправно. Но недавно стал плохо замораживать продукты в морозилке.
Мне понравилось, что у вас мастер приехал в позднее вечернее время, не пришлось отпрашиваться специально с работы.
Раиса Алексеевна
Хочу выразить благодарность мастеру Виктору.
Очень вежливый внимательный и судя по результату проведенной работы – профессионал своего дела.
Побольше таких бы людей в наш мир. Всем добра!
Дмитрий Александрович
Хочу сказать огромное спасибо за качественно проделанную работу. До этого случая сталкивался тоже с ремонтом холодильника, так сказать есть с чем сравнивать.
23.09. 2020
2020Михаил
Нам от старых хозяев квартиры остался холодильник Атлант. Сказали, что не ремонтируемый. С 7 этажа вытаскивать некому. Позвонил ребятам, приехал мастер. Пришлось им конечно повозиться с ремонтом, но теперь у нас 2 холодильника. Так, что не спешите расстраиваться, а обратитесь к мастерам своего дела.
22.08.2020Ирина Владимировна
Добрый день.
Приезжал мастер Андрей, приятный и внимательный молодой человек.
Быстро и аккуратно исправил неисправность, за что большое спасибо!
Masha_Kruglova
Хочу сказать спасибо мастеру Роману, который отремонтировал мой холодильник Горенье. Был внимателен и вежлив – видно, что воспитан и профессионал своего дела. Побольше бы таких людей в наш мир! С уважением Мария Сергеевна К. П.С.
09.06.2020Виктория
Отдельное спасибо диспетчеру, по телефону выслушал не перебивая, задавал уточняющие вопросы и если так можно выразится поставил диагноз поломки и озвучил полную стоимость работ.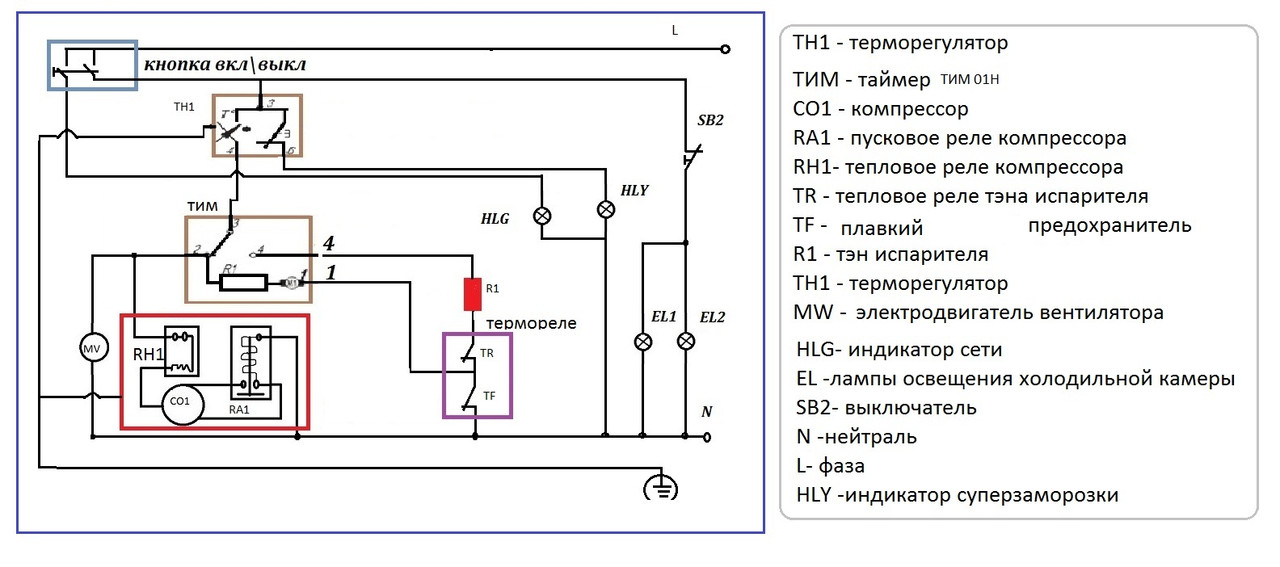 Пришедший мастер после диагностики подтвердил предположения поломки и стоимость ремонта. Все прошло хорошо,, холодильник заработал, мастер оставил гарантийный талон.
Пришедший мастер после диагностики подтвердил предположения поломки и стоимость ремонта. Все прошло хорошо,, холодильник заработал, мастер оставил гарантийный талон.
Владимир Ефимович
Хочу оставить отзыв о работе ребят. Заявку принял приятный внимательный СПЕЦИАЛИСТ, а не диспетчер. Мастер приехал в назначенное время, отремонтировал все сразу же на месте, чему очень рада (стоимость ремонта по факту соответствовала заявленной стоимости по телефону). РЕКОМЕНДУЮ
29.05.2020Евгений
Было приятно иметь дело с ребятами из Сервис цента холод. Все качественно и без лишних наворотов.
Спасибо.
RITA
Добрый день. Хочу оставить отзыв о работе сотрудника Евгения и всей фирме в целом.
Все оперативно и качественно. Сам мастер Евгений видно, что профессионал и знает свое дело. Желаю вам профессионального успеха! Маргарита.
Олег Иванович C.
Рекомендую эту фирму: 1. Хорошо общаются по телефону (все понятно и внятно) 2. Приехали не опоздав! – плюс ко всему в удобное для меня время – мне не пришлось менять свои планы 3. Стоимость работ была озвучена предварительно по телефону и в процессе ремонта НЕ увеличилась! Мастер был аккуратен к моему дому и был вежлив. Спасибо!
Приехали не опоздав! – плюс ко всему в удобное для меня время – мне не пришлось менять свои планы 3. Стоимость работ была озвучена предварительно по телефону и в процессе ремонта НЕ увеличилась! Мастер был аккуратен к моему дому и был вежлив. Спасибо!
Холодильник вестфрост, ул. Мосфильмовская 31 А
Владимир
приезжал мастер 12 января.. несмотря на то, что изначально была озвучена одна стоимость работ (предварительно по телефону) в конце был удевлен стоимость оказалась меньше… как пояснил мастер вышел из строя менее дорогой узел, для меня это было странно, что объявляли одну стоимость а взяли меньше.. в наше время такое мне не встречалось)))
12.01.2020ГРАНИТ Мемори
Обращался к ребятам по вопросу ремонта моего холодильника. Всё как говориться по делу и ничего лишнего. Ценовая политика меня устроила.
Отдельное спасибо мастеру Евгению.
Таймер, електронний таймер, тим-01, тим-01н, принцип роботи, схема живлення, підключення, перевірка,
Електронний таймер ТІМ- 01 (застосовується для управління оттайкой випарника холодильників No Frost.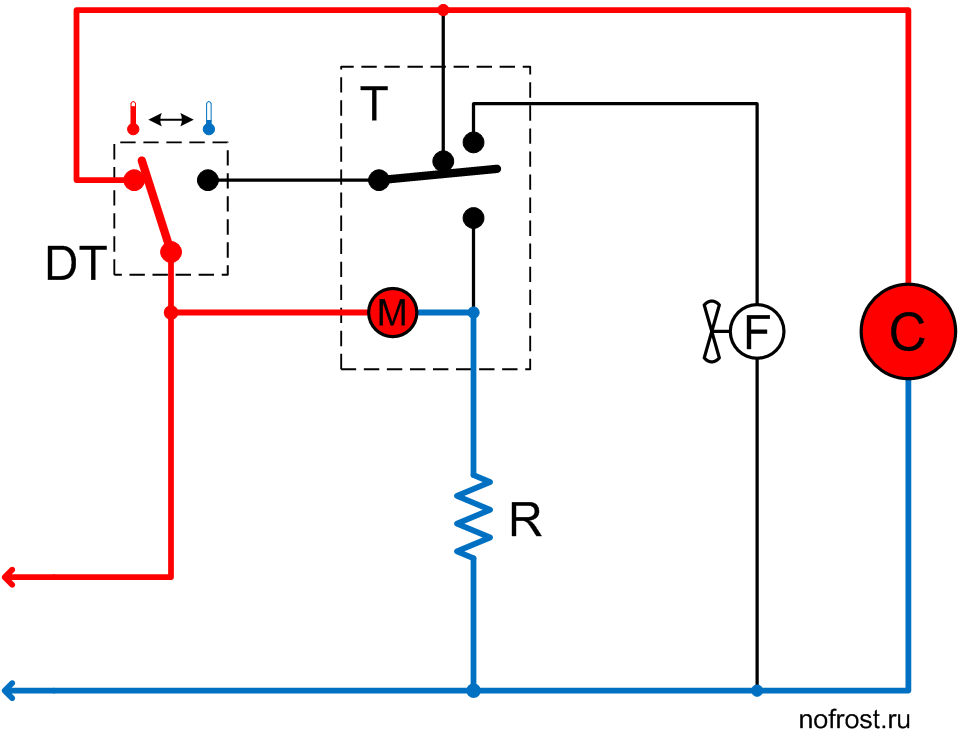 Full No Frost. Total No Frost – Індезіт, Арістон
Full No Frost. Total No Frost – Індезіт, Арістон
Цикл оттайкі – через 8, 12 годин роботи компресора (час стоянки не враховується)
Відтайка починається за умови – температура в морозильній камері не вище – 10 ° С
Закінчення оттайкі – при температурі на термореле +10 С. Час відтаювання залежить від кількості інею на випарнику.
Електронний таймер ТІМ- 01Н
При першому включенні холодильника, при досягненні температури в морозильній камері -10 С, таймер входить в режим розморожування – використовується для контролю роботи системи відтаювання.
Відтайка починається за умови – температура в морозильній камері не вище – 10 ° С
Закінчення оттайкі – при температурі на термореле + 10С. Час відтаювання залежить від кількості інею на випарнику
Параметри для перевірки:
– ручна установка режиму розмерзання
– включення холодильного режиму
Перевіряють в наступній послідовності
1. При замкнутих контактах теплового реле, коли температура в морозильній камері нижче – 8 (+/- 5 С) (для реле ТАБ -Т) або – 10 (+ \ – 3) (для COMBI-100 b 261N), натискають кнопку таймера.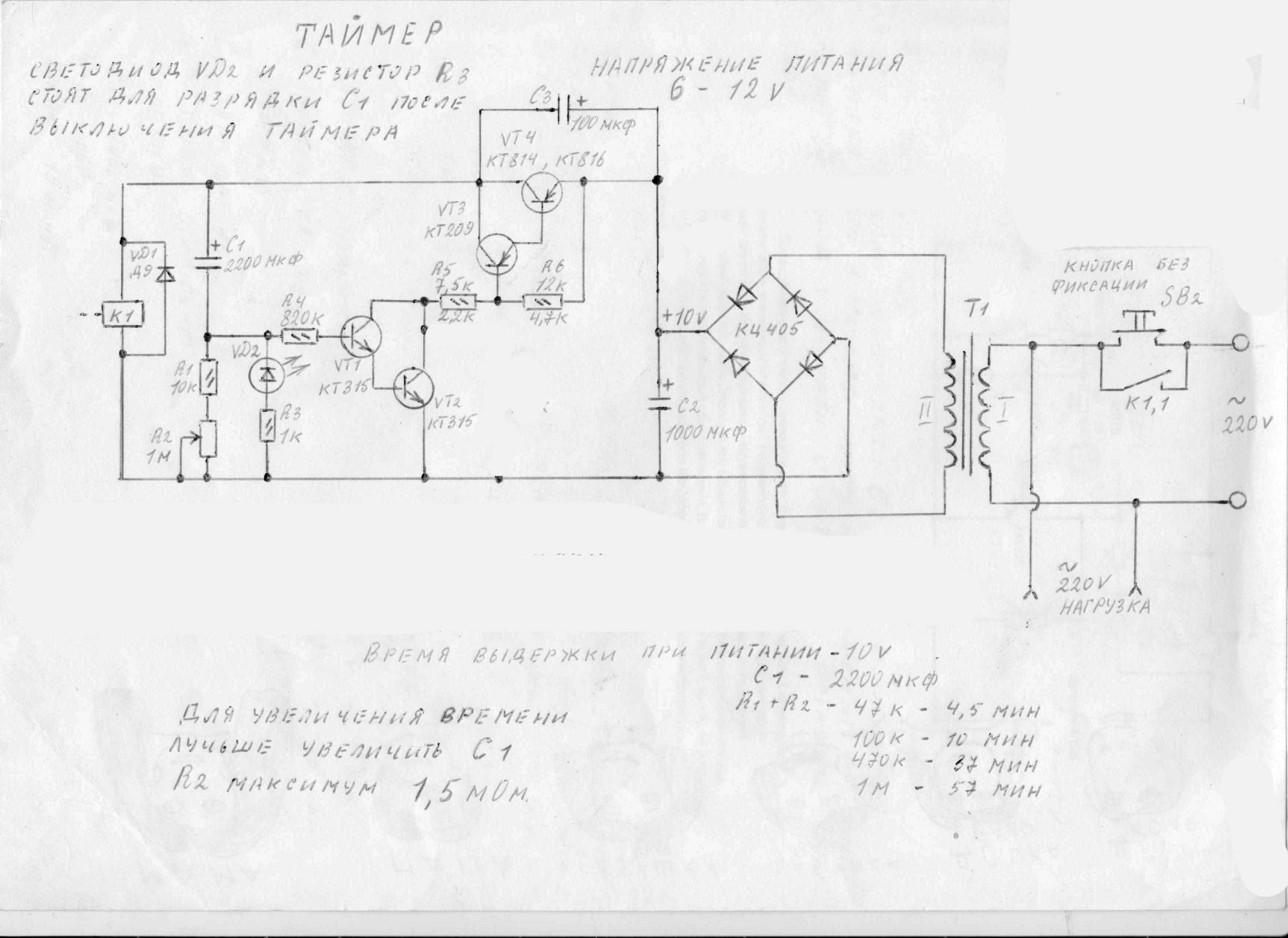 При цьому таймер повинен перевести систему «NO FROST» в режим відтаювання (відключається компресор і включаються тени).
При цьому таймер повинен перевести систему «NO FROST» в режим відтаювання (відключається компресор і включаються тени).
2. Отсоединяют провід термозапобіжника від комутаційної колодки, тим самим імітується розмикання контактів теплового реле, після цього якщо використовується таймер ТІМ -01 – компресор включається через 7 (+ \ – 3) хв.
3. Після цього відновлюють цілісність схеми холодильника. При необхідності замінюють дефектний таймер
Індезіт NBA18FNF Індезіт C132NFQ.016
ТЕУ-01-2 – оттайка електронагрівачем, збереження інформації про режим роботи при відключенні електроживлення на протязі 8 годин, тривалість режиму активної оттайкі визначається тепловим реле, але не більше.
РВ-1 електронний таймер призначений для автоматичного управління режимом оттайкі
Холодильники з системою «NO FROST» відрізняються від холодильників із статичним охолодженням тим, що в морозильній камері у них немає статичного випарника, на полиці, якого поміщаються продукти. До складу системи входить таймер, випарник, вентилятор, тен оттайкі і система зливу талої води. Випарник системи «NO FROST»
До складу системи входить таймер, випарник, вентилятор, тен оттайкі і система зливу талої води. Випарник системи «NO FROST»
Нажмите здесь, чтобы купить этот продукт в Tactical Online
Нажмите на изображение ниже, чтобы загрузить технический паспорт
Ниже приведены примеры подключения / конфигурации для приложений TIM-01:
| ||||||||||||||||

Объяснение простых схем таймера задержки
В этом посте мы обсудим создание простых таймеров задержки с использованием очень обычных компонентов, таких как транзисторы, конденсаторы и диоды. Все эти схемы будут производить задержку включения или задержки выключения с интервалами времени на выходе на заранее определенный период, от нескольких секунд до многих минут. Все конструкции полностью регулируются.
Все эти схемы будут производить задержку включения или задержки выключения с интервалами времени на выходе на заранее определенный период, от нескольких секунд до многих минут. Все конструкции полностью регулируются.
Важность таймеров задержки
Во многих приложениях электронных схем задержка в несколько секунд или минут становится решающим требованием для обеспечения правильной работы схемы. Без указанной задержки схема может выйти из строя или даже выйти из строя.
Давайте подробно разберем различные конфигурации.
Вы также можете прочитать о таймерах задержки на основе IC 555. Рекомендуется для вас!
Использование одиночного транзистора и кнопки
Первая принципиальная схема показывает, как транзисторы и несколько других пассивных компонентов могут быть подключены для получения заданных выходов времени задержки.
Транзистор снабжен обычным базовым резистором для функций ограничения тока.
Светодиод, который используется здесь только для индикации, ведет себя как нагрузка коллектора схемы.
Конденсатор, который является важной частью схемы, занимает определенное положение в схеме, мы видим, что он размещен на другом конце базового резистора, а не непосредственно на базе транзистора.
Кнопка используется для включения цепи.
При кратковременном нажатии кнопки положительное напряжение от линии питания поступает на базовый резистор и включает транзистор, а затем светодиод.
Однако в ходе вышеуказанного действия конденсатор также полностью заряжается.
При отпускании кнопки, хотя питание базы отключается, транзистор продолжает работать с помощью накопленной энергии в конденсаторе, который теперь начинает разряжать накопленный заряд через транзистор.
Светодиод также остается включенным, пока конденсатор полностью не разрядится.
Те значение конденсатора определяет время задержки или время, в течение которого транзистор остается в проводящем режиме.
Наряду с конденсатором, номинал базового резистора также играет важную роль в определении времени, в течение которого транзистор остается включенным после отпускания кнопки.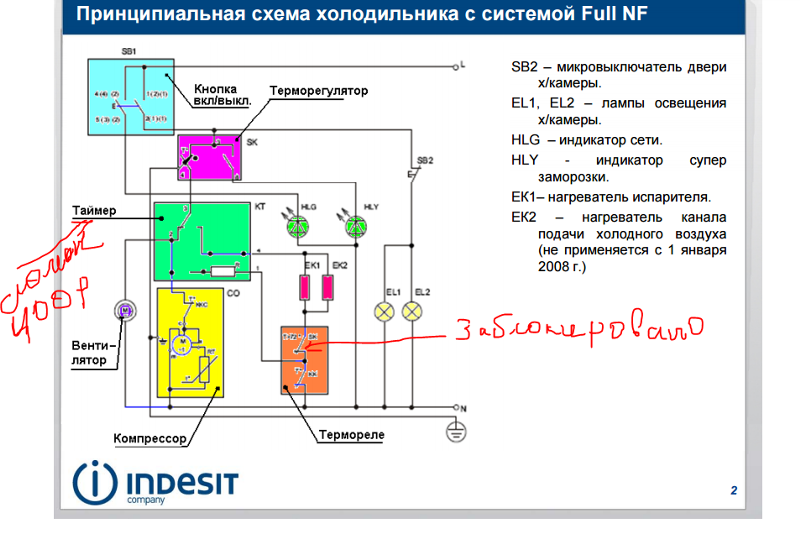
Однако схема, использующая только один транзистор, сможет создавать задержки, которые могут составлять всего несколько секунд.
При добавлении еще одного транзисторного каскада (следующий рисунок) указанный выше диапазон времени задержки может быть значительно увеличен.
Добавление еще одного транзисторного каскада увеличивает чувствительность схемы, что позволяет использовать более высокие значения резистора синхронизации, тем самым увеличивая диапазон временной задержки схемы.
Дизайн печатной платы
Видео демонстрация
 youtube.com/embed/iwlGt2cOTVY?feature=oembed” frameborder=”0″ allow=”accelerometer; autoplay; clipboard-write; encrypted-media; gyroscope; picture-in-picture” allowfullscreen=””/>
youtube.com/embed/iwlGt2cOTVY?feature=oembed” frameborder=”0″ allow=”accelerometer; autoplay; clipboard-write; encrypted-media; gyroscope; picture-in-picture” allowfullscreen=””/>
Использование симистора:
На следующем изображении показано, как указанная выше схема таймера задержки может быть интегрирована с симистором и использоваться для переключения нагрузки сети переменного тока
Вышеупомянутое может быть дополнительно модифицировано с помощью автономного силового бестрансформаторного источника питания, как показано ниже:
Без кнопки
Если вышеуказанная конструкция предназначена для использования без кнопки, то же самое может быть реализовано, как показано на следующей диаграмме:
Вышеупомянутый эффект задержки выключения без нажатия кнопки может быть дополнительно улучшен путем использования двух транзисторов NPN и использования конденсатора между базой / землей левого NPN
Следующая схема показывает, как соответствующая кнопка может стать неактивной, как только она будет нажата, и пока таймер задержки находится в активированном состоянии.
В это время любое дальнейшее нажатие кнопки не влияет на таймер, пока выход активен или пока таймер не завершит свою операцию задержки.
Задержка от внешнего триггера
Проблема, заданная г-ном Гленом (одним из преданных читателей этого блога):
У меня есть ситуация, когда у меня есть импульс 12 В, который длится около 4 секунд (от поворотного переключателя, вращается медленным двигателем), но мне нужно всего лишь полсекунды (чтобы вызвать механический звонок / перезвон).
Есть ли способ взять длинный импульс в цепь и послать намного более короткий импульс?
Решение вышеуказанной проблемы представлено на следующей схеме:
Двухшаговый последовательный таймер
Вышеупомянутая схема может быть изменена для создания двухступенчатого последовательного генератора задержки. Эта схема была запрошена одним из заядлых читателей этого блога, г-ном Марко.
Простая цепь аварийной сигнализации отключения с задержкой показана на следующей диаграмме.
Схема запрошена Dmats.
Следующая схема была запрошена Fastshack3
Таймер задержки с реле
«Я ищу схему, которая будет управлять выходным реле. Это будет сделано на 12 В, а последовательность будет инициирована ручным переключателем.
Мне понадобится регулируемая задержка времени (возможно, отображаемое время) после отпускания переключателя, тогда выход будет включаться в течение настраиваемого времени (также возможно отображается) перед отключением.
Последовательность не будет перезапущена, пока не будет нажата кнопка и снова выпустили.
Время после отпускания кнопки составляет от 250 миллисекунд до 5 секунд. Время включения выхода для включения реле составляет от 500 миллисекунд до 30 секунд. Дайте мне знать, если вы можете что-то поделать. Спасибо! »
До сих пор мы научились делать простые таймеры задержки выключения, теперь давайте посмотрим, как мы можем построить простую схему таймера задержки включения, которая позволяет подключенной нагрузке на выходе включаться с некоторой заранее заданной задержкой после выключения питания. ВКЛ.
ВКЛ.
Объясненная схема может использоваться для всех приложений, в которых требуется функция начальной задержки включения для подключенной нагрузки после включения сетевого питания.
Схема работы схемы таймера задержки включения
Показанная диаграмма довольно проста, но очень впечатляюще предоставляет необходимые действия, кроме того, период задержки является переменным, что делает установку чрезвычайно полезной для предлагаемых приложений.
Функционирование можно понять по следующим пунктам:
Предполагая, что нагрузка, требующая задержки включения, подключена к контактам реле, при включении питания 12 В постоянного тока проходит через R2, но не может достигнуть базы T1, потому что изначально C2 действует как короткое замыкание на землю.
Таким образом, напряжение проходит через R2, падает до соответствующих пределов и начинает заряжать C2.
Как только C2 заряжается до уровня, который развивает потенциал от 0,3 до 0,6 В (+ стабилитрон) на базе T1, T1 мгновенно включается, переключая T2, а затем реле .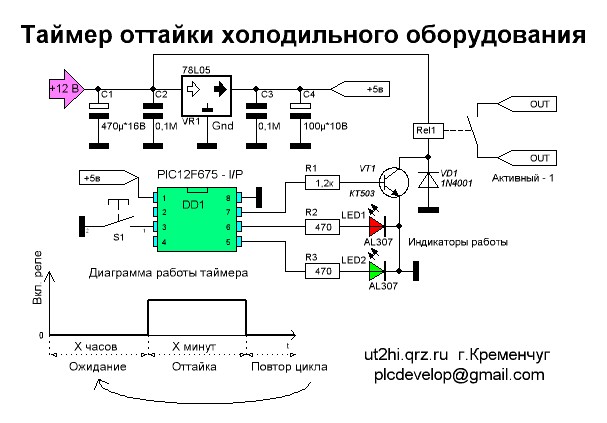 … наконец, нагрузка становится тоже включен.
… наконец, нагрузка становится тоже включен.
Вышеупомянутый процесс вызывает необходимую задержку для включения нагрузки.
Период задержки может быть установлен соответствующим выбором значений R2 и C2.
R1 гарантирует, что C2 быстро разряжается через него, так что схема достигает положения ожидания как можно скорее.
D3 блокирует заряд от базы T1.
Список деталей
R1 = 1o0K (резистор для разряда C2, когда цепь выключена))
R2 = 330K (синхронизирующий резистор)
R3 = 10K
R4 = 10K
D1 = стабилитрон 3 В (опционально, можно заменить на провод)
D2 = 1N4007
D3 = 1N4148
T1 = BC547
T2 = BC557
C2 = 33 мкФ / 25 В (синхронизирующий конденсатор)
Реле = SPDT, 12 В / 400 Ом
PCB Design
Примечание по применению
узнайте, как приведенная выше схема таймера задержки включения становится применимой для решения следующей проблемы, представленной одним из ярых последователей этого блога, г-ном. Нишант.
Нишант.
Проблема цепи:
Здравствуйте, сэр,
У меня есть автоматический стабилизатор напряжения на 1 кВА. У него есть один недостаток: когда он включен, очень высокое напряжение выдается в течение примерно 1,5 с (поэтому CFL и лампочка часто перегорают) после что напряжение становится нормальным.
Я открыл стабилизатор, он состоит из автотрансформатора, 4 реле на 24 В, каждое реле подключено к отдельной цепи (каждое из
10K предустановок, BC547, стабилитрон, BDX53BFP npn, пара транзисторов Дарлингтона IC, конденсатор 220 мкФ / 63 В. , Конденсатор 100uF / 40V, 4 диода и несколько резисторов).
Эти схемы питаются от понижающего трансформатора, и выходной сигнал этих схем берется через соответствующий конденсатор 100 мкФ / 40 В. и подается на соответствующее реле. Что делать для решения проблемы. Пожалуйста, помогите мне. Нарисованная вручную принципиальная схема прилагается. .
Решение проблемы цепи
Проблема в приведенной выше схеме может быть вызвана двумя причинами: одно из реле на мгновение включается, соединяя неправильные контакты с выходом, или одно из ответственных реле стабилизируется с правильным напряжением. через некоторое время после включения питания.
через некоторое время после включения питания.
Поскольку имеется более одного реле, выявление неисправности и ее устранение может быть немного утомительным … Схема таймера задержки включения, описанная в вышеупомянутой статье, может быть действительно очень эффективной для обсуждаемой цели.
Подключения достаточно простые.
Используя 7812 IC, таймер задержки может питаться от существующего источника питания 24 В стабилизатора.
Затем замыкающие контакты реле задержки могут быть подключены последовательно с проводкой выходного разъема стабилизатора.
Вышеупомянутая проводка мгновенно решила бы проблемы, так как теперь выход будет переключаться через некоторое время во время включения питания, давая достаточно времени для внутренних реле, чтобы установить правильные напряжения на их выходных контактах.
Отзыв от г-на Билла
Привет Свагатам,
Я наткнулся на вашу страницу, исследуя Интернет, чтобы сделать мою задержку более последовательной.![]() Сначала немного справочной информации.
Сначала немного справочной информации.
Я занимаюсь драг-рейсингом и запускаю машину при первом взгляде на 3-ю янтарную лампочку, когда рождественская елка спускается.
Я использую выключатель транс-тормоза, который нажат, чтобы заблокировать автоматическую коробку передач одновременно переднего и заднего хода.
Это позволяет увеличить обороты двигателя для увеличения мощности для запуска. Когда кнопка отпускается, трансмиссия выключается с заднего хода и движется вперед на высоких оборотах.
Это все равно, что выскакивать сцепление на автомобиле с механической коробкой передач, в любом случае моя машина реагирует на это быстро, и в результате появляется красный свет, уезжает слишком рано, и вы проигрываете гонку.
Уменьшение времени реакции на запуск – это все, и это игра на сотни тысяч с большими мальчиками, поэтому я поставил переключатель транс-тормоза на реле и поместил комбо на 1100 мкФ на реле, чтобы задержать его запуск.
Из-за автомобильной электроники я не верю, что есть точное напряжение, заряжающее эту крышку каждый раз, когда я активирую эту схему, и точность является ключевой, поэтому я купил стабилизатор мощности на Ebay, который потребляет 8-15 вольт и дает постоянный 12вольт на выходе.
Это перевернуло мой сезон, но я считаю, что эту схему можно было бы сделать более точной и более легким способом варьировать время задержки, а не заменять комбо.
Также я должен установить диод перед реле, а не сейчас, потому что все, что есть, это выключатель – куда пойдет ток? Я ни в коем случае не инженер-электрик, но у меня есть некоторые знания по устранению неисправностей в аудио высокого класса в течение многих лет.
Хотел бы получить ваши мысли – спасибо
Билл Кореки
Анализ и решение схемы
Привет, Билл,
Я приложил схему регулируемой цепи задержки, пожалуйста, проверьте ее. Вы можете использовать его для указанной цели.
Предустановка 100K может использоваться и настраиваться для получения точных коротких периодов задержки в соответствии с вашими требованиями.
Тем не менее, обратите внимание, что для правильной работы реле на 12 В напряжение питания должно быть минимум 11 В, если это не выполняется, цепь может работать неправильно.
С уважением.
Простой таймер задержки от 5 до 20 минут
В следующем разделе обсуждается простая схема таймера задержки от 5 до 20 минут для конкретного промышленного применения.
Идею предложил мистер Джонатан.
Технические требования
Пытаясь найти решение моей проблемы в Google, я наткнулся на вашу публикацию выше.
Я пытаюсь понять, как построить лучший контроллер Sous Vide.Основная проблема заключается в том, что моя водяная баня имеет очень высокий гистерезис, и при нагреве от более низких температур температура будет превышать примерно 7 градусов по сравнению с температурой, при которой прекращается питание.
Он также очень хорошо изолирован, с зазором между внутренним и внешним резервуаром, который заставляет его действовать как термос, из-за чего требуется очень много времени, чтобы спуститься от любого превышения температуры. Мой ПИД-регулятор имеет контрольный выход SSR и релейный выход аварийной сигнализации.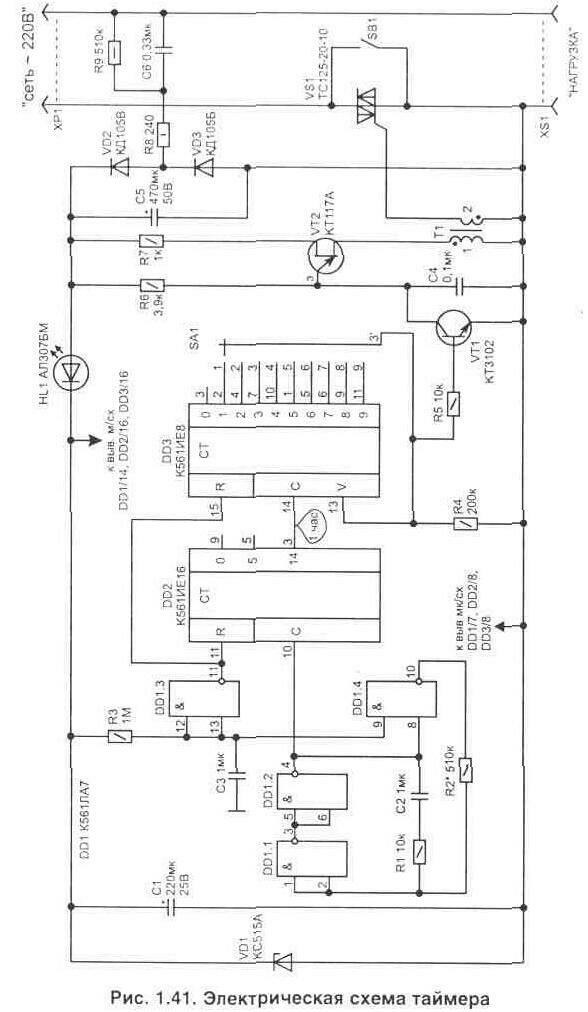
Аварийный сигнал можно запрограммировать как аварийный сигнал ниже предела со смещением от заданного значения.Я могу использовать источник питания на пять вольт, который у меня уже есть, для моего циркуляционного двигателя, чтобы он работал через реле аварийной сигнализации и управлял тем же SSR, что и управляющий выход.
Затем я установлю будильник, чтобы он оставался включенным, пока температура не поднимется выше заданного значения минус 7 градусов. Это позволит регулировать настройку ПИД-регулятора без учета начального повышения температуры.
Поскольку я знаю, что последние несколько градусов будут достигнуты без какой-либо подачи питания, мне бы очень хотелось отложить любое распознавание управляющего сигнала примерно на пять минут после отключения будильника, так как он все равно будет звонить для тепла.
Это та часть, для которой мне еще предстоит разобраться в схеме. Я имею в виду нормально замкнутое реле, включенное последовательно с управляющим выходом, которое удерживается разомкнутым сигналом тревоги.
Когда сигнал тревоги прекращается, мне нужна задержка порядка пяти минут, прежде чем реле вернется в свое нормально замкнутое состояние «выключено».
Я был бы признателен за помощь с задержкой отключения части схемы реле. Мне нравится простота начального дизайна на странице, но у меня такое впечатление, что с ними не справиться и около пяти минут.
Спасибо,
Джонатан Лундквист
Схема схемы
Следующая схема простой схемы таймера задержки от 5 до 20 минут может быть подходящим образом применена для указанного выше приложения.
Схема использует IC4049 для необходимых вентилей НЕ, которые сконфигурированы как компараторы напряжения.
5 ворот, включенных параллельно, образуют чувствительную секцию и обеспечивают триггер с требуемой временной задержкой для последующих каскадов буфера и драйвера реле.
Управляющий вход поступает от выхода тревоги, как указано в приведенном выше описании. Этот вход становится коммутационным напряжением для предлагаемой схемы таймера.
Этот вход становится коммутационным напряжением для предлагаемой схемы таймера.
При получении этого триггера вход 5 вентилей НЕ изначально удерживается на логическом нуле, потому что конденсатор заземляет начальный триггер через потенциометр 2 м2.
В зависимости от настройки 2м2 конденсатор начинает заряжаться, и в тот момент, когда напряжение на конденсаторе достигает распознаваемого значения, вентили НЕ возвращают свой выход на низкий логический уровень, который преобразуется как высокий логический уровень на выходе правого сингла. НЕ ворота.
Это мгновенно запускает подключенный транзистор и реле для требуемой задержки выхода через контакты реле.
Потолок 2M2 можно отрегулировать для определения требуемых задержек.
Принципиальная схема
Ограниченное по времени кормление продлевает продолжительность жизни дрозофилы в зависимости от периферийных часов
Abstract
Ограниченное по времени кормление / прием пищи (TRF / TRE) – ограничение не количества пищи, а дневного временного окна потребление пищи – это диетическое вмешательство, которое, как было показано, улучшает маркеры здоровья у модельных организмов и людей, но неясно, приводят ли эти преимущества к положительному воздействию на старение и долголетие. Мы демонстрируем здесь, что TRF надежно продлевает продолжительность жизни в короткоживущем генетически поддающемся модельному организму Drosophila melanogaster . Среднее увеличение продолжительности жизни TRF колеблется от ~ 10% до ~ 50% в зависимости от пола, репродуктивного статуса, продолжительности TRF и генотипа. Положительный эффект TRF на долголетие не зависит от приема пищи и, по крайней мере, частично зависит от функционирующих циркадных часов: преимущества TRF для долголетия отменяются у аритмических мутантов per 0 и tim 01 мутантов. как при постоянном освещении, предполагая, что кормление по времени действует как zeitgeber, разделяя прием пищи и связанные с ней метаболические процессы на определенные фазы дня и ночи.TRF-опосредованное увеличение продолжительности жизни не затрагивается у мух, чьи нервные циркадные часы были отменены генетически, указывая на периферические часы как на цель TRF, опосредовавшую продление продолжительности жизни.
Мы демонстрируем здесь, что TRF надежно продлевает продолжительность жизни в короткоживущем генетически поддающемся модельному организму Drosophila melanogaster . Среднее увеличение продолжительности жизни TRF колеблется от ~ 10% до ~ 50% в зависимости от пола, репродуктивного статуса, продолжительности TRF и генотипа. Положительный эффект TRF на долголетие не зависит от приема пищи и, по крайней мере, частично зависит от функционирующих циркадных часов: преимущества TRF для долголетия отменяются у аритмических мутантов per 0 и tim 01 мутантов. как при постоянном освещении, предполагая, что кормление по времени действует как zeitgeber, разделяя прием пищи и связанные с ней метаболические процессы на определенные фазы дня и ночи.TRF-опосредованное увеличение продолжительности жизни не затрагивается у мух, чьи нервные циркадные часы были отменены генетически, указывая на периферические часы как на цель TRF, опосредовавшую продление продолжительности жизни.
Введение
Старение, ухудшение функции организма с течением времени (Flatt and Schmidt, 2009), наблюдалось почти у всех протестированных форм жизни (Ackermann et al., 2003; Lewis and Buffenstein, 2016) и проявляется в молекулярный, клеточный, тканевый и организменный уровни.Исследователи определили несколько коррелятов старения, включая геномную нестабильность, истощение теломер, эпигенетические изменения, потерю протеостаза, нарушение чувствительности к питательным веществам, митохондриальную дисфункцию, клеточное старение, истощение стволовых клеток и нарушение межклеточной коммуникации (López-Otín et al., 2013). Причины этих изменений не ясны, но процессы-кандидаты включают макромолекулярное повреждение, вызванное реактивными формами кислорода (АФК), временные и стохастические изменения уровней молекул, регулирующих долголетие, ухудшение циркадных часов и гипертрофию (Gems and Partridge, 2013; Kenyon, 2010; Mattis, Sehgal, 2016; Musiek, Holtzman, 2016).Относительная важность как причин, так и признаков старения и их взаимосвязи друг с другом остается неясной.
Старение контролируется как внутренними факторами, так и факторами окружающей среды, включая диету. Впервые обнаруженный у крыс во время Великой депрессии, ученые обнаружили, что сокращение доступности пищи, как ни странно, не сокращает, а увеличивает продолжительность жизни (McCay et al., 1989). Позже было обнаружено, что ограничение питания (DR) продлевает продолжительность жизни у многих видов, от дрожжей до приматов, и улучшает маркеры здоровья у людей, что указывает на эволюционно законсервированный процесс (Chaix et al., 2019b; Колман и др., 2009; Партридж и др., 2005; Walker et al., 2005). Предварительный генетический скрининг, исследующий генетику старения, показал, что действительно необходимы два взаимосвязанных пути определения питательных веществ, путь передачи сигналов инсулин / инсулиноподобного фактора роста 1 (IGF-1) (IIS) и пути-мишени рапамицина (TOR). о влиянии DR на долголетие (Kenyon, 2010). DR, по крайней мере частично, действует на пути IIS и TOR, изменяя клеточные программы транскрипции, с участием киназы S6 и факторов транскрипции FOXO. Последующие события включают активацию транскрипционной программы, делающую клетки более устойчивыми к молекулярным повреждениям и ограничивающие рост. В дополнение к DR, включая ограничение калорийности и ограничение потребления определенных питательных веществ или аминокислот, было показано, что другие диетические вмешательства оказывают положительное влияние на старение: прерывистое голодание, которое включает в себя различные протоколы кормления или приема пищи и голодание более 24 часов (Паттерсон и Sears, 2017), и кормление с ограничением по времени (TRF), при котором потребление пищи ограничено ежедневно определенными часами дня, а общее диетическое потребление – нет (Chaix et al., 2019б). У Drosophila было показано, что TRF ослабляет возрастное снижение сердечной деятельности во время старения (Gill et al., 2015), а также улучшает сон и мышечную функцию (Villanueva et al., 2019). У мышей TRF снижает уровень холестерина и сахара в крови, снижает массу тела, воспаление и дисбактериоз, одновременно увеличивая расход энергии, контроль моторики и выносливость (Chaix et al.
Последующие события включают активацию транскрипционной программы, делающую клетки более устойчивыми к молекулярным повреждениям и ограничивающие рост. В дополнение к DR, включая ограничение калорийности и ограничение потребления определенных питательных веществ или аминокислот, было показано, что другие диетические вмешательства оказывают положительное влияние на старение: прерывистое голодание, которое включает в себя различные протоколы кормления или приема пищи и голодание более 24 часов (Паттерсон и Sears, 2017), и кормление с ограничением по времени (TRF), при котором потребление пищи ограничено ежедневно определенными часами дня, а общее диетическое потребление – нет (Chaix et al., 2019б). У Drosophila было показано, что TRF ослабляет возрастное снижение сердечной деятельности во время старения (Gill et al., 2015), а также улучшает сон и мышечную функцию (Villanueva et al., 2019). У мышей TRF снижает уровень холестерина и сахара в крови, снижает массу тела, воспаление и дисбактериоз, одновременно увеличивая расход энергии, контроль моторики и выносливость (Chaix et al. , 2019b). Исследования на людях подтвердили потенциал TRE как способствующего укреплению здоровья вмешательства и показали, что TRF может помочь улучшить ряд метаболических маркеров, включая вес, жировые отложения, артериальное давление, потребление энергии и выносливость, особенно у людей с риском метаболического синдрома (Chaix et al. ., 2019b; Wilkinson et al., 2020). TRF отличается от DR, поскольку его польза для здоровья, похоже, не требует снижения калорийности (Gill et al., 2015; Hatori et al., 2012; Mitchell et al., 2019). Механизм действия TRF не совсем понятен, однако он, по-видимому, увеличивает амплитуду циркадных ритмов различных физиологических параметров, которые обычно сглаживаются во время старения, представляя возможный механизм действия (Manoogian and Panda, 2017).
, 2019b). Исследования на людях подтвердили потенциал TRE как способствующего укреплению здоровья вмешательства и показали, что TRF может помочь улучшить ряд метаболических маркеров, включая вес, жировые отложения, артериальное давление, потребление энергии и выносливость, особенно у людей с риском метаболического синдрома (Chaix et al. ., 2019b; Wilkinson et al., 2020). TRF отличается от DR, поскольку его польза для здоровья, похоже, не требует снижения калорийности (Gill et al., 2015; Hatori et al., 2012; Mitchell et al., 2019). Механизм действия TRF не совсем понятен, однако он, по-видимому, увеличивает амплитуду циркадных ритмов различных физиологических параметров, которые обычно сглаживаются во время старения, представляя возможный механизм действия (Manoogian and Panda, 2017).
Хотя влияние TRF на здоровье и старение описано, в настоящее время неизвестно, увеличивает ли TRF продолжительность жизни (Francesco et al., 2018), хотя мыши, получавшие однократный прием пищи, а также мыши с ограничением калорийности, демонстрируют увеличение продолжительности жизни, которое коррелирует с добровольным ежедневным голоданием (Acosta-Rodríguez et al. , 2017; Mitchell et al., 2019). Поскольку эффекты TRF, как и другие диетические вмешательства, по-видимому, сохраняются в ходе эволюции, для решения этого вопроса полезно использовать короткоживущие генетически поддающиеся лечению модельные организмы. В настоящем исследовании мы использовали Drosophila melanogaster , которые живут всего три месяца, что дает нам уникальную возможность провести многомерный многопараметрический анализ, чтобы выяснить, влияет ли TRF на продолжительность жизни и каковы соответствующие биологические факторы и лежащие в основе механизмы. .
, 2017; Mitchell et al., 2019). Поскольку эффекты TRF, как и другие диетические вмешательства, по-видимому, сохраняются в ходе эволюции, для решения этого вопроса полезно использовать короткоживущие генетически поддающиеся лечению модельные организмы. В настоящем исследовании мы использовали Drosophila melanogaster , которые живут всего три месяца, что дает нам уникальную возможность провести многомерный многопараметрический анализ, чтобы выяснить, влияет ли TRF на продолжительность жизни и каковы соответствующие биологические факторы и лежащие в основе механизмы. .
Результаты
TRF продлевает продолжительность жизни у
Drosophila Чтобы оценить, влияет ли TRF на продолжительность жизни, мы подвергали самок, самцов и девственницу Drosophila от изогенного штамма дикого типа к суточному 12-часовому питанию, 12-часовому режиму агара в регулярные циклы свет-темнота (LD) на протяжении всей их жизни (рис. 1A), позволяя мухам есть в течение дня и иметь доступ к содержащему воду агару в течение ночи, чтобы избежать высыхания. Поскольку плотность популяции, генетический фон и состав пищи независимо друг от друга влияют на продолжительность жизни (Иванов и др., 2015; Джоши и Мюллер, 1997; Skorupa et al., 2008), каждый из этих параметров оставался постоянным во всех экспериментах, если явно не указано иное (более подробную информацию см. В разделе «Материалы и методы»). TRF значительно увеличил среднюю продолжительность жизни у спарившихся самок мух (далее называемых «мухами», если не указано иное) в среднем с 49 до 60 дней (+ 18%), но не у спарившихся самцов или девственных самок (рис. 1B, C, Таблица 1 и Дополнение Рис. 1). Было показано, что краткосрочные TRF / TRE полезны для здоровья сердца и сна у Drosophila (Gill et al., 2015) и метаболических маркеров у крыс и человека (de Goede et al., 2019; Hutchison et al., 2019; Jamshed et al., 2019; Ravussin et al., 2019). Чтобы определить, влияет ли TRF короче, чем продолжительность жизни, на продолжительность жизни, мы протестировали 10, 20, 30, 40 и 50 дней TRF по сравнению с ALF.
Поскольку плотность популяции, генетический фон и состав пищи независимо друг от друга влияют на продолжительность жизни (Иванов и др., 2015; Джоши и Мюллер, 1997; Skorupa et al., 2008), каждый из этих параметров оставался постоянным во всех экспериментах, если явно не указано иное (более подробную информацию см. В разделе «Материалы и методы»). TRF значительно увеличил среднюю продолжительность жизни у спарившихся самок мух (далее называемых «мухами», если не указано иное) в среднем с 49 до 60 дней (+ 18%), но не у спарившихся самцов или девственных самок (рис. 1B, C, Таблица 1 и Дополнение Рис. 1). Было показано, что краткосрочные TRF / TRE полезны для здоровья сердца и сна у Drosophila (Gill et al., 2015) и метаболических маркеров у крыс и человека (de Goede et al., 2019; Hutchison et al., 2019; Jamshed et al., 2019; Ravussin et al., 2019). Чтобы определить, влияет ли TRF короче, чем продолжительность жизни, на продолжительность жизни, мы протестировали 10, 20, 30, 40 и 50 дней TRF по сравнению с ALF. TRF продемонстрировал доза-ответный эффект в зависимости от продолжительности TRF с минимальной продолжительностью 20 дней для достижения увеличения продолжительности жизни (фиг. 1D, левая панель и таблица 1). Чтобы оценить, является ли TRF по-прежнему полезным во второй половине жизни, мы подвергали мух воздействию TRF в возрасте от 30 до 60 дней.TRF во второй половине жизни мух значительно продлил продолжительность жизни (рис. 1D, правая панель), показывая, что TRF полезен независимо от возраста, в котором она началась.
TRF продемонстрировал доза-ответный эффект в зависимости от продолжительности TRF с минимальной продолжительностью 20 дней для достижения увеличения продолжительности жизни (фиг. 1D, левая панель и таблица 1). Чтобы оценить, является ли TRF по-прежнему полезным во второй половине жизни, мы подвергали мух воздействию TRF в возрасте от 30 до 60 дней.TRF во второй половине жизни мух значительно продлил продолжительность жизни (рис. 1D, правая панель), показывая, что TRF полезен независимо от возраста, в котором она началась.
Ограниченное по времени кормление (TRF) продлевает продолжительность жизни самок дикого типа Drosophila . Показаны отдельные эксперименты, обобщенные на рис. 1С. Мух переносили в zeitgeber time (ZT) 12 из пищи в агар и ZT 0 из агара в пищу, чтобы ограничить потребление пищи до 12 часов в течение дня (группа TRF) или из пищи в пищу в качестве контроля (ALF).Выживаемость мух оценивали ежедневно. TRF значительно увеличивает продолжительность жизни у спарившихся самок (A), но не у спарившихся самцов (B). (A) Графики выживаемости для TRF на самках. n (ALF, TRF) = (82,84), (116,105), (79,67), (59,69), (153,142), (95,75) соответственно. (B) Графики выживаемости для TRF у самцов. n (ALF, TRF) = (34,38), (95,90), (77,83) соответственно.
(A) Графики выживаемости для TRF на самках. n (ALF, TRF) = (82,84), (116,105), (79,67), (59,69), (153,142), (95,75) соответственно. (B) Графики выживаемости для TRF у самцов. n (ALF, TRF) = (34,38), (95,90), (77,83) соответственно.
Сводка описанных эффектов TRF. Левый столбец обозначает экспериментальные группы, разъясняющие роль различных параметров в эффектах TRF. Гистограмма показывает положительное (зеленый), нейтральный (черный) или отрицательный (красный) эффект TRF на долголетие.Значения P, выделенные жирным шрифтом справа, указывают на значительные изменения.
Рис. 1. Ограниченное по времени кормление (TRF) продлевает продолжительность жизни самок дикого типа Drosophila . (A) Упрощенная схема используемого метода TRF. Мух переносили в zeitgeber time (ZT) 12 из пищи в агар и ZT 0 из агара в пищу, чтобы ограничить потребление пищи до 12 часов в течение дня (группа TRF) или из пищи в пищу в качестве контроля (ALF). Выживаемость мух оценивали ежедневно. (B) TRF значительно увеличивает продолжительность жизни у спарившихся самок, но не у спарившихся самцов или девственниц (ALF n = 63,80,87, TRF n = 81,83,89, соответственно).(C) TRF воспроизводимо увеличивает продолжительность жизни у (слева) спарившихся самок (ALF ns = 63, 59, 79, 95, 116, 82, 153. TRF ns = 81, 69, 67, 75, 105, 84, 142), но не (справа) самцы (ALF ns = 80, 34, 77, 95, TRF ns = 83, 38, 83, 90) в разных экспериментах (см. дополнительные графики выживаемости на рис. 1). * = P≤0,05 (D) TRF действует в зависимости от продолжительности. (Слева) Спарившиеся самки подвергались TRF в течение 10, 20, 30, 40, 50 дней (ALF n = 86, 10, 20, 30, 40, 50 дней, TRF ns = 84, 75, 77, 79, 82 соответственно) или (Справа) в возрасте от 30 до 60 дней (ALF n = 84, TRF n = 84).(Слева) Увеличение продолжительности жизни с помощью TRF наблюдается при минимальной продолжительности 20 дней и увеличивается с более длительным TRF. (Справа) TRF во второй половине жизни мух значительно увеличивает продолжительность жизни.
(B) TRF значительно увеличивает продолжительность жизни у спарившихся самок, но не у спарившихся самцов или девственниц (ALF n = 63,80,87, TRF n = 81,83,89, соответственно).(C) TRF воспроизводимо увеличивает продолжительность жизни у (слева) спарившихся самок (ALF ns = 63, 59, 79, 95, 116, 82, 153. TRF ns = 81, 69, 67, 75, 105, 84, 142), но не (справа) самцы (ALF ns = 80, 34, 77, 95, TRF ns = 83, 38, 83, 90) в разных экспериментах (см. дополнительные графики выживаемости на рис. 1). * = P≤0,05 (D) TRF действует в зависимости от продолжительности. (Слева) Спарившиеся самки подвергались TRF в течение 10, 20, 30, 40, 50 дней (ALF n = 86, 10, 20, 30, 40, 50 дней, TRF ns = 84, 75, 77, 79, 82 соответственно) или (Справа) в возрасте от 30 до 60 дней (ALF n = 84, TRF n = 84).(Слева) Увеличение продолжительности жизни с помощью TRF наблюдается при минимальной продолжительности 20 дней и увеличивается с более длительным TRF. (Справа) TRF во второй половине жизни мух значительно увеличивает продолжительность жизни.
Частичное восстановление нормальной продолжительности жизни с помощью TRF у мутантов с коротким сном
Drosophila На продолжительность жизни влияет ряд факторов, включая температуру, световые условия, генетический фон и сон (Bushey et al., 2010; Lamb, 1968). ; Малик и Кидвелл, 1966; Шиба и др., 2000). Большинство мутантов сна Drosophila демонстрируют укороченную продолжительность жизни (Koh et al., 2008; Kume et al., 2005; Liu et al., 2014; Rogulja and Young, 2012; Stavropoulos and Young, 2011). Чтобы проверить, ограничиваются ли эффекты TRF только нормально живущими мухами дикого типа или же они способствуют долголетию мутаций, влияющих на сон и, в большинстве случаев, на продолжительность жизни, мы выявили короткоживущих мутантов сна sss P1 , inc. 1 , wakeD1, wakeD2 , а также фумин , который был описан как имеющий нормальный срок службы для TRF или ALF на всю жизнь. TRF значительно увеличил продолжительность жизни у всех протестированных самок-мутантов-самок (рис. 2А и таблица 1), с увеличением продолжительности жизни на 50% для sss P1 , 14% для wakeD2 , 13% для inc 1 , 12% для wakeD1 и 9% для фумин . Интересно, что долголетию у некоторых мутантов мужского сна также способствовал TRF (рис. 2C, таблица 1), в отличие от мух дикого типа, у которых TRF не увеличивал продолжительность жизни самцов (рис.1Б, таблица 1, доп. Рис. 1Б). Эти данные показывают, что эффекты TRF не ограничиваются нормально живущими мухами дикого типа, но способствуют продолжительности жизни в различной степени у других генотипов, особенно у мутантов с коротким сном. Поскольку TRF работает с некоторыми штаммами самцов, мы пришли к выводу, что в принципе возможно продлить продолжительность жизни как самок, так и самцов, используя TRF. Увеличение продолжительности сна, по-видимому, не требуется для влияния TRF на долголетие: хотя мы подтвердили ранее сообщаемое увеличение общего суточного сна у мужчин, вызванное TRF (Gill et al.
TRF значительно увеличил продолжительность жизни у всех протестированных самок-мутантов-самок (рис. 2А и таблица 1), с увеличением продолжительности жизни на 50% для sss P1 , 14% для wakeD2 , 13% для inc 1 , 12% для wakeD1 и 9% для фумин . Интересно, что долголетию у некоторых мутантов мужского сна также способствовал TRF (рис. 2C, таблица 1), в отличие от мух дикого типа, у которых TRF не увеличивал продолжительность жизни самцов (рис.1Б, таблица 1, доп. Рис. 1Б). Эти данные показывают, что эффекты TRF не ограничиваются нормально живущими мухами дикого типа, но способствуют продолжительности жизни в различной степени у других генотипов, особенно у мутантов с коротким сном. Поскольку TRF работает с некоторыми штаммами самцов, мы пришли к выводу, что в принципе возможно продлить продолжительность жизни как самок, так и самцов, используя TRF. Увеличение продолжительности сна, по-видимому, не требуется для влияния TRF на долголетие: хотя мы подтвердили ранее сообщаемое увеличение общего суточного сна у мужчин, вызванное TRF (Gill et al. , 2015), данные не показаны), у этих мух не наблюдается увеличения продолжительности жизни (Рис. 1B, Supp. Fig. 1B). Напротив, самки мух, получавших TRF, не показывают изменений в общем дневном сне, хотя их дневной сон увеличен по сравнению с мухами с ALF (дополнительный рис. 2). Важно отметить, что эффекты TRF на сон отсутствуют у мутанта сна inc 1 (Supp. Fig. 2), хотя эффекты долголетия отсутствуют (Fig. 2), и, наоборот, на 0 мутанты показывают усиливается ночной сон (Supp.Рис. 2), но не влияет на долголетие (Рис. 4), предполагая, что TRF увеличивает продолжительность жизни независимо от воздействия на сон.
, 2015), данные не показаны), у этих мух не наблюдается увеличения продолжительности жизни (Рис. 1B, Supp. Fig. 1B). Напротив, самки мух, получавших TRF, не показывают изменений в общем дневном сне, хотя их дневной сон увеличен по сравнению с мухами с ALF (дополнительный рис. 2). Важно отметить, что эффекты TRF на сон отсутствуют у мутанта сна inc 1 (Supp. Fig. 2), хотя эффекты долголетия отсутствуют (Fig. 2), и, наоборот, на 0 мутанты показывают усиливается ночной сон (Supp.Рис. 2), но не влияет на долголетие (Рис. 4), предполагая, что TRF увеличивает продолжительность жизни независимо от воздействия на сон.
TRF и сон. Самки дикого типа, inc 1 , per 0 и tim 01 мух лечили TRF или ALF. Сон оценивали через 14, 30 или 45 дней. (A) Для всех графиков сна показано среднее значение сна за 24 часа за 4 дня, объединенное в интервал сна за 30 минут. (B) Количественная оценка (A).Показаны гистограммы среднего общего дневного сна, дневного сна и ночного сна, а также SEM. Для каждого условия было протестировано 16 мух. Для оценки значимости использовались t-критерии. * = P≤0,05
(B) Количественная оценка (A).Показаны гистограммы среднего общего дневного сна, дневного сна и ночного сна, а также SEM. Для каждого условия было протестировано 16 мух. Для оценки значимости использовались t-критерии. * = P≤0,05
TRF увеличивает продолжительность жизни у мутантов сна с укороченной продолжительностью жизни в зависимости от часов, пола и генотипа. Самок или самцов мух содержали на TRF или ALF на протяжении всей их жизни. (A) Все указанные генотипы демонстрируют TRF-опосредованное увеличение продолжительности жизни. Inc 1 (ALF n = 91, TRF n = 87), sss P1 (ALF n = 58, TRF n = 50), fmn (ALF n = 136, TRF n = 113), wakeD1 (ALF n = 144, TRF n = 149) и wakeD2 (ALF n = 151, TRF n = 123).(B) tim 01 нулевая мутация обращает TRF-опосредованное увеличение продолжительности жизни у inc 1 мух (ALF n = 87, TRF n = 91). (C) TRF способствует увеличению продолжительности жизни в inc 1 (ALF n = 88, TRF n = 86) и wakeD1 (ALF n = 155, TRF n = 127), но не wakeD2 (ALF n = 63, TRF n = 84), fmn (ALF n = 114, TRF n = 120) и sss P1 (ALF n = 57, TRF n = 54) летает.
(C) TRF способствует увеличению продолжительности жизни в inc 1 (ALF n = 88, TRF n = 86) и wakeD1 (ALF n = 155, TRF n = 127), но не wakeD2 (ALF n = 63, TRF n = 84), fmn (ALF n = 114, TRF n = 120) и sss P1 (ALF n = 57, TRF n = 54) летает.
Мутации часов обращают опосредованное TRF увеличение продолжительности жизни светозависимым образом. Самки дикого типа, на 0 или tim 01 мух содержались на 12:12 TRF или ALF в течение всей их жизни. (A) В 12:12 свет: темнота (LD) мутации дикого типа (слева), per (в центре) и tim (справа) (ALF n = 116,83,78, TRF n = 105,60,95, соответственно ) отменить TRF-опосредованное увеличение продолжительности жизни (слева). (B) При постоянном освещении TRF больше не увеличивает продолжительность жизни у мух дикого типа, но увеличивает продолжительность жизни у мух на 0 и tim 01 (ALF n = 135,119,137, TRF n = 107, 71,104 соответственно) мутантов. (C) В постоянной темноте TRF способствует увеличению продолжительности жизни у дикого типа, на 0 и на 01 (ALF n = 150 147 150, TRF n = 143 125 151, соответственно).
(C) В постоянной темноте TRF способствует увеличению продолжительности жизни у дикого типа, на 0 и на 01 (ALF n = 150 147 150, TRF n = 143 125 151, соответственно).
Влияние TRF на кормление и массу тела. (A) Самка или (B) самец дикого типа, по 0 и tim 01 мухи прошли 14 дней TRF или ALF в 12:12 LD. Потребление пищи измеряли в группах из 10 мух в течение 12 часов (группа TRF) или 24 часов (группа ALF) с использованием анализа капиллярного питателя (CAFE).Каждая точка соответствует потреблению пищи 10 мухами. (C) Самка или (D) самец дикого типа, на 0 и tim 01 мухи подверглись 14-дневной TRF или ALF в 12:12 LD и были взвешены в группах по 10 человек. в ZT 10. (E) Самка или (F) самец inc 1 , wakeD2 и fmn подверглись 30-дневной TRF или ALF в 12:12 LD и были взвешены в группах по 10 человек в ZT. 10. Каждая точка представляет собой вес 10 мух, разделенный на 10.Среднее значение и SEM указаны красным. Тест Манна-Уитни оценил статистическую значимость. * = P≤0,05
10. Каждая точка представляет собой вес 10 мух, разделенный на 10.Среднее значение и SEM указаны красным. Тест Манна-Уитни оценил статистическую значимость. * = P≤0,05
Эффекты TRF зависят от часов и света
Циркадные часы состоят из молекулярных механизмов, присутствующих в большинстве клеток и сохраняемых в процессе эволюции, что позволяет организмам согласовывать физиологические параметры с ежедневными циклами окружающей среды, обеспечивая временную организацию биологических процессов и поведения (Патке и др., 2019). Было показано, что влияние TRF на здоровье сердца зависит от циркадных часов (Gill et al., 2015). Чтобы выяснить, необходимы ли интактные молекулярные циркадные часы также для TRF-опосредованного увеличения продолжительности жизни, мы протестировали аритмические на 0 и tim 01 часовых мутантов (Konopka and Benzer, 1971; Sehgal et al. др., 1994). Влияние TRF на продолжительность жизни отсутствует у мутантов per 0 и tim 01 (рис.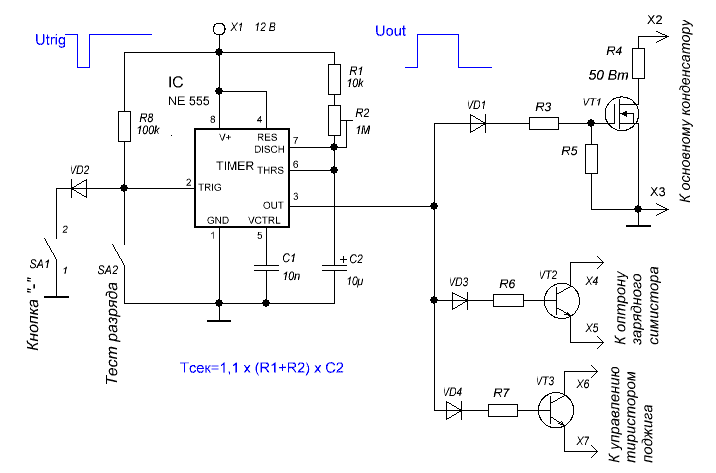 3A), а также у мутантов inc 1 , tim 905 01 двойной мутант (рис.2B), предполагая, что функционирующие циркадные часы необходимы для TRF-опосредованных эффектов долголетия. Этот вывод подтверждается экспериментами TRF, проведенными при постоянном освещении (LL), которые вызывают у мух дикого типа аритмию из-за постоянной переустановки молекулярных часов из-за вневременной деградации (Myers et al., 1996; Zeng et al., 1996) . В LL TRF имеет противоположный эффект по сравнению с LD: он вреден для долголетия мух (рис. 3B, слева). В постоянной темноте преимущества TRF для мух дикого типа сохраняются (рис.3C), а TRF также был полезен для часовых мутантов по 0 и tim 01 , у которых продолжительность жизни увеличивалась также при постоянном освещении (рис. 3B). Эти данные предполагают, что эффекты TRF зависят от функционирующих циркадных часов, а также от цикла свет / темнота.
3A), а также у мутантов inc 1 , tim 905 01 двойной мутант (рис.2B), предполагая, что функционирующие циркадные часы необходимы для TRF-опосредованных эффектов долголетия. Этот вывод подтверждается экспериментами TRF, проведенными при постоянном освещении (LL), которые вызывают у мух дикого типа аритмию из-за постоянной переустановки молекулярных часов из-за вневременной деградации (Myers et al., 1996; Zeng et al., 1996) . В LL TRF имеет противоположный эффект по сравнению с LD: он вреден для долголетия мух (рис. 3B, слева). В постоянной темноте преимущества TRF для мух дикого типа сохраняются (рис.3C), а TRF также был полезен для часовых мутантов по 0 и tim 01 , у которых продолжительность жизни увеличивалась также при постоянном освещении (рис. 3B). Эти данные предполагают, что эффекты TRF зависят от функционирующих циркадных часов, а также от цикла свет / темнота.
Преимущества продолжительности жизни TRF не зависят от снижения потребления пищи
Ограничение калорийности (или диетического питания) (DR) – известный механизм увеличения продолжительности жизни у различных модельных организмов (Colman et al., 2009; Партридж и др., 2005). Чтобы проверить, приводит ли TRF просто к DR, мы измерили потребление пищи мухами после 14 или 32 дней TRF или ALF. Используя анализ капиллярного питателя (CAFE (Ja et al., 2007)), мы определили, что TRF не заставляет мух есть меньше, поскольку обработанные TRF самки мух дикого типа, которые демонстрируют увеличение продолжительности жизни при TRF, потребляют такое же количество пищи во время Через 12 часов у них есть доступ к пище, чем у контрольных животных ad libitum за 24 часа (фиг. 4A, дополнительная фиг. 3A), демонстрируя, что TRF отличается от DR.В отличие от этого, tim 01 мух действительно демонстрируют снижение потребления пищи при TRF (рис. 4A), однако это не приводит к увеличению продолжительности жизни в нашем анализе (рис. 3A). Чтобы проверить, как TRF влияет на массу тела мух, которая связана с различными маркерами метаболизма, а также со старением у Drosophila и людей (Fontana and Hu, 2014; Gáliková and Klepsatel, 2018), мы взвешивали мух в 3 или 4 группах. из 10 через 14, 30 или 45 дней ежедневного применения TRF или ALF.Хотя самки мух дикого типа демонстрируют потерю веса после 45 дней приема TRF (см. Рис. 3B), 14 или 30 дней приема TRF не оказывают значительного влияния на их вес (рис. 4C, E). Потеря веса также наблюдалась для некоторых других генотипов ( на 0 , wakeD2 , рис. 3 C, E, Supp. Рис. 3B) и для самцов дикого типа после 30 дней TRF (рис. 3F), однако эти состояния не связаны с увеличением продолжительности жизни, опосредованным TRF. Следовательно, потеря веса происходила независимо от TRF-индуцированных эффектов долголетия, что позволяет предположить, что потеря веса не является основным механизмом, ведущим к увеличению продолжительности жизни.
3A). Чтобы проверить, как TRF влияет на массу тела мух, которая связана с различными маркерами метаболизма, а также со старением у Drosophila и людей (Fontana and Hu, 2014; Gáliková and Klepsatel, 2018), мы взвешивали мух в 3 или 4 группах. из 10 через 14, 30 или 45 дней ежедневного применения TRF или ALF.Хотя самки мух дикого типа демонстрируют потерю веса после 45 дней приема TRF (см. Рис. 3B), 14 или 30 дней приема TRF не оказывают значительного влияния на их вес (рис. 4C, E). Потеря веса также наблюдалась для некоторых других генотипов ( на 0 , wakeD2 , рис. 3 C, E, Supp. Рис. 3B) и для самцов дикого типа после 30 дней TRF (рис. 3F), однако эти состояния не связаны с увеличением продолжительности жизни, опосредованным TRF. Следовательно, потеря веса происходила независимо от TRF-индуцированных эффектов долголетия, что позволяет предположить, что потеря веса не является основным механизмом, ведущим к увеличению продолжительности жизни.
Влияние TRF на кормление. (A) Самки мух дикого типа прошли 32 дня TRF или ALF за 12:12 LD. Потребление пищи измеряли в группах из 10 мух в течение 12 часов (группа TRF) или 24 часов (группа ALF) с использованием анализа капиллярного питателя (CAFE). Каждая точка соответствует потреблению пищи 10 мухами. (B) Вес самки, на 0 и tim 01 мухи подверглись 45 дням TRF или ALF в 12:12 LD и были взвешены группами по 10 человек в ZT 10.Каждая точка представляет собой вес 10 мух, разделенный на 10. Среднее значение и SEM показаны красным. Тест Манна-Уитни оценил статистическую значимость. * = P≤0,05
TRF-опосредованное увеличение продолжительности жизни не требует нейронных циркадных часов
В то время как большинство клеток имеют молекулярные циркадные часы, периферические ткани, включая мышцы, печень и почки, могут получать информацию о времени суток либо непосредственно из сигналов окружающей среды, либо из определенной структуры мозга, называемой центральными или главными часами (Patke et al. , 2019). Drosophila Локомоторные ритмы и цикл сна / бодрствования контролируются так называемыми часовыми нейронами в центральном мозге, часть которых, наряду с фактором диспергирования пигмента циркадных нейропептидов, также участвует в старении (Donlea et al., 2014; Умезаки и др., 2012). Чтобы выяснить, требуются ли функциональные нейронные часы для воздействия TRF на долголетие, мы использовали пан-нейрональный драйвер Gal4 nsyb-Gal4 , чтобы сбить tim- РНК с помощью UAS-tim-RNAi , что приводит к поведенческой аритмичности мух. (Прилож.Рис.4). В то время как повсеместно распространенный tim-РНКи во всех тканях с использованием тубулина-Gal4, фенокопирует отмену эффектов TRF, наблюдаемых у нуль-мутантов tim 01 (рис. 4A, правая панель и 5A), нейрон-специфичных РНКи оставили эффекты TRF нетронутыми (рис. 5B), предполагая, что основанные на нервной системе циркадные часы незаменимы для эффектов TRF на продолжительность жизни и что периферические часы могут поддерживать TRF-опосредованное увеличение продолжительности жизни.
, 2019). Drosophila Локомоторные ритмы и цикл сна / бодрствования контролируются так называемыми часовыми нейронами в центральном мозге, часть которых, наряду с фактором диспергирования пигмента циркадных нейропептидов, также участвует в старении (Donlea et al., 2014; Умезаки и др., 2012). Чтобы выяснить, требуются ли функциональные нейронные часы для воздействия TRF на долголетие, мы использовали пан-нейрональный драйвер Gal4 nsyb-Gal4 , чтобы сбить tim- РНК с помощью UAS-tim-RNAi , что приводит к поведенческой аритмичности мух. (Прилож.Рис.4). В то время как повсеместно распространенный tim-РНКи во всех тканях с использованием тубулина-Gal4, фенокопирует отмену эффектов TRF, наблюдаемых у нуль-мутантов tim 01 (рис. 4A, правая панель и 5A), нейрон-специфичных РНКи оставили эффекты TRF нетронутыми (рис. 5B), предполагая, что основанные на нервной системе циркадные часы незаменимы для эффектов TRF на продолжительность жизни и что периферические часы могут поддерживать TRF-опосредованное увеличение продолжительности жизни.
Свободная деятельность мух с нокдауном tim в разных тканях.2-3-дневных мух указанных генотипов вовлекали в циклы LD на 3 дня и переводили в DD для оценки ритмичности. Повсеместная tim-RNAi с использованием tub-Gal4 , а также нокдаун нейронов с использованием nsyb-Gal4 вызывают аритмичность, в то время как гетерозиготные контроли остаются ритмичными. Показаны репрезентативные примеры подсчета активности отдельных мух на двойном графике. В отличие от самцов мух, у спариваемых нами самок Drosophila всех генотипов наблюдается более высокая доля аритмичных мух, что может быть связано с сокращением сна и другими ранее описанными половыми диморфизмами в локомоторном поведении Drosophila (см. Обсуждение).
Рис. 5. TRF продлевает срок службы независимо от нейронных часов. (A) Повсеместный нокдаун tim с использованием tim-RNAi отменяет TRF-опосредованное увеличение продолжительности жизни (слева: нокдаун (ALF n = 75, TRF n = 43), в центре: tub-Gal4 , управление драйвером (ALF n = 80, TRF n = 70), справа: tim – UAS-RNAi контроль (ALF n = 75,80,67, TRF n = 24)). (B) Слева: пан-нейрональная tim-RNAi (ALF n = 81, TRF n = 36) не влияет на TRF-опосредованное увеличение продолжительности жизни.В центре: nsyb-Gal4 управление нейрональным драйвером (ALF n = 77, TRF n = 53)
(B) Слева: пан-нейрональная tim-RNAi (ALF n = 81, TRF n = 36) не влияет на TRF-опосредованное увеличение продолжительности жизни.В центре: nsyb-Gal4 управление нейрональным драйвером (ALF n = 77, TRF n = 53)
Обсуждение
TRF / TRE привлекает внимание в последние годы как диетическое вмешательство, приносящее пользу для здоровья людей и животных, но TRF влияние на долголетие, а также основные принципы преимуществ TRF остаются в значительной степени неизвестными. В настоящем исследовании мы демонстрируем увеличение продолжительности жизни с помощью TRF, зависящее от часов и света, но не зависящее от нейронов.
TRF как новый механизм продления жизни
Ограничение диеты (DR) – сокращение потребления калорий или определенных макроэлементов – было описано как мощный метод продления продолжительности жизни дрожжей, C.elegans, Drosophila , мышей и приматов (Colman et al., 2009; Partridge et al., 2005). Мы наблюдаем, что преимущества продолжительности жизни, опосредованные TRF, не зависят от снижения потребления пищи, что согласуется с предыдущими исследованиями на мухах и мышах, показавшими, что другие преимущества для здоровья, вызванные TRF, также не зависят от DR (Gill et al. , 2015; Hatori et al., 2012; Mitchell et al., 2019). Хотя потеря веса происходит в некоторых условиях, когда TRF продлевает продолжительность жизни, она также происходит, когда продолжительность жизни не увеличивается, и наоборот, что позволяет предположить, что потеря веса не является основной причиной увеличения продолжительности жизни TRF, а скорее является следствием его (пока неизвестного) механизма.В отличие от DR, TRF демонстрирует зависящие от продолжительности преимущества, начиная с двух недель TRF (рис. 1D), в то время как эффекты DR на уровень смертности, как было показано, сохраняются только через 48 часов (Mair et al., 2003). У мышей DR, по-видимому, работает только у молодых животных (Hahn et al., 2019), тогда как в настоящем исследовании TRF одинаково эффективен у молодых и старых животных. Эти данные добавляют дополнительные доказательства того, что TRF механически отличается от DR. С другой стороны, в некоторых экспериментальных парадигмах DR трудно отличить от TRF.В двух недавних исследованиях была предпринята попытка сравнить TRF и DR у мышей, однако мыши с DR самостоятельно наложили TRF и съели все пищевые гранулы в узком интервале от одного до пяти часов, что фактически сделало DR неотличимым от TRF (Acosta-Rodríguez et al.
, 2015; Hatori et al., 2012; Mitchell et al., 2019). Хотя потеря веса происходит в некоторых условиях, когда TRF продлевает продолжительность жизни, она также происходит, когда продолжительность жизни не увеличивается, и наоборот, что позволяет предположить, что потеря веса не является основной причиной увеличения продолжительности жизни TRF, а скорее является следствием его (пока неизвестного) механизма.В отличие от DR, TRF демонстрирует зависящие от продолжительности преимущества, начиная с двух недель TRF (рис. 1D), в то время как эффекты DR на уровень смертности, как было показано, сохраняются только через 48 часов (Mair et al., 2003). У мышей DR, по-видимому, работает только у молодых животных (Hahn et al., 2019), тогда как в настоящем исследовании TRF одинаково эффективен у молодых и старых животных. Эти данные добавляют дополнительные доказательства того, что TRF механически отличается от DR. С другой стороны, в некоторых экспериментальных парадигмах DR трудно отличить от TRF.В двух недавних исследованиях была предпринята попытка сравнить TRF и DR у мышей, однако мыши с DR самостоятельно наложили TRF и съели все пищевые гранулы в узком интервале от одного до пяти часов, что фактически сделало DR неотличимым от TRF (Acosta-Rodríguez et al. , 2017; Mitchell et al., 2019). Новый анализ распределения пищи по времени позволит исследователям на мышах различать эти два диетических вмешательства (J. Takahashi, неопубликовано). Еще одно диетическое вмешательство, способствующее пользе для здоровья у различных модельных организмов, включая C.elegans, Drosophila , мышей, а у людей – периодическое голодание (IF), при котором организмы подвергаются длительным (> = 24 часам) периодам голодания. Хотя было показано, что IF улучшает метаболические маркеры (Patterson and Sears, 2017) и продолжительность жизни, действуя независимо от пути TOR (Catterson et al., 2018) – основной мишени DR -, неясно, действует ли TRF через то же самое. молекулярные пути как IF. В будущем будет интересно лучше разобраться в механистических различиях и сходствах между различными диетическими вмешательствами, направленными на старение.
, 2017; Mitchell et al., 2019). Новый анализ распределения пищи по времени позволит исследователям на мышах различать эти два диетических вмешательства (J. Takahashi, неопубликовано). Еще одно диетическое вмешательство, способствующее пользе для здоровья у различных модельных организмов, включая C.elegans, Drosophila , мышей, а у людей – периодическое голодание (IF), при котором организмы подвергаются длительным (> = 24 часам) периодам голодания. Хотя было показано, что IF улучшает метаболические маркеры (Patterson and Sears, 2017) и продолжительность жизни, действуя независимо от пути TOR (Catterson et al., 2018) – основной мишени DR -, неясно, действует ли TRF через то же самое. молекулярные пути как IF. В будущем будет интересно лучше разобраться в механистических различиях и сходствах между различными диетическими вмешательствами, направленными на старение.
Половые различия в эффектах TRF
Мы видим, что эффекты TRF на продолжительность жизни более универсальны для генотипов у женщин, чем у мужчин.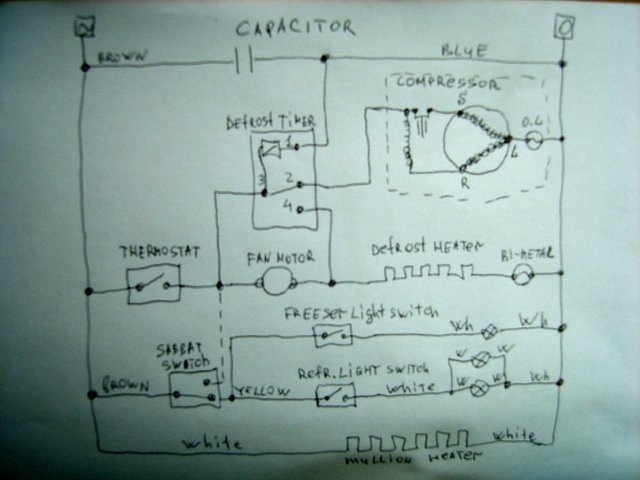 TRF увеличил среднюю продолжительность жизни у всех протестированных генотипов самок, в то время как у самцов три различных мутантных линии, но не мух дикого типа, показали значительные улучшения с нашим анализом (рис. 2 и таблица 1). Хотя исторически в большинстве областей биологических исследований, включая старение, использовались только мужчины, половые различия становятся важной областью исследований сами по себе.Мужские и женские особи и модельные организмы, включая Drosophila , различаются по возрасту, а самки обычно живут дольше, однако эта взаимосвязь не является универсальной и полностью понятной и сильно зависит от факторов, включая генетику, репродуктивный статус и окружающую среду, включая диету (Austad and Fischer , 2016). Репродуктивный статус, в дополнение к общей продолжительности жизни, по-видимому, также влияет на динамику выживаемости на уровне популяции. Дева, похоже, следуют иной траектории выживания, чем самцы и спарившиеся самки (рис.1 B), при отсутствии задержки гибели мух на ранней стадии жизни.
TRF увеличил среднюю продолжительность жизни у всех протестированных генотипов самок, в то время как у самцов три различных мутантных линии, но не мух дикого типа, показали значительные улучшения с нашим анализом (рис. 2 и таблица 1). Хотя исторически в большинстве областей биологических исследований, включая старение, использовались только мужчины, половые различия становятся важной областью исследований сами по себе.Мужские и женские особи и модельные организмы, включая Drosophila , различаются по возрасту, а самки обычно живут дольше, однако эта взаимосвязь не является универсальной и полностью понятной и сильно зависит от факторов, включая генетику, репродуктивный статус и окружающую среду, включая диету (Austad and Fischer , 2016). Репродуктивный статус, в дополнение к общей продолжительности жизни, по-видимому, также влияет на динамику выживаемости на уровне популяции. Дева, похоже, следуют иной траектории выживания, чем самцы и спарившиеся самки (рис.1 B), при отсутствии задержки гибели мух на ранней стадии жизни. Это отклонение от классического «S-образного» графика выживаемости, по-видимому, характерно для девственной Drosophila , поскольку предыдущие исследования также показали аналогичную динамику выживаемости (Markow, 2011; Villanueva et al., 2019). Было показано, что у мух TRF благотворно влияет на сон у мужчин ((Gill et al., 2015), данные по самкам в этом исследовании не приводятся) и улучшает здоровье сердца и мышечную функцию у обоих полов (Gill et al., 2015; Вильянуэва и др., 2019). Предыдущая работа, изучающая влияние TRF на продолжительность жизни у обоих полов, не обнаружила никаких преимуществ, в отличие от данных, представленных здесь. Разница, вероятно, связана с репродуктивным статусом используемых мух: хотя в предыдущем исследовании, по-видимому, использовались девственные самцы и самки, мы обнаружили, что у самок дикого типа только спарившиеся самки реагируют увеличением продолжительности жизни на лечение TRF, состояние не тестировалось. в предыдущем исследовании (Villanueva et al.
Это отклонение от классического «S-образного» графика выживаемости, по-видимому, характерно для девственной Drosophila , поскольку предыдущие исследования также показали аналогичную динамику выживаемости (Markow, 2011; Villanueva et al., 2019). Было показано, что у мух TRF благотворно влияет на сон у мужчин ((Gill et al., 2015), данные по самкам в этом исследовании не приводятся) и улучшает здоровье сердца и мышечную функцию у обоих полов (Gill et al., 2015; Вильянуэва и др., 2019). Предыдущая работа, изучающая влияние TRF на продолжительность жизни у обоих полов, не обнаружила никаких преимуществ, в отличие от данных, представленных здесь. Разница, вероятно, связана с репродуктивным статусом используемых мух: хотя в предыдущем исследовании, по-видимому, использовались девственные самцы и самки, мы обнаружили, что у самок дикого типа только спарившиеся самки реагируют увеличением продолжительности жизни на лечение TRF, состояние не тестировалось. в предыдущем исследовании (Villanueva et al. , 2019). Хотя мы не знаем, почему только спарившиеся самки показывают увеличение продолжительности жизни с TRF, различные соображения могут играть роль в дифференциальной реакции спарившихся самок.Самки мух крупнее самцов, демонстрируют различия в метаболизме и менее чувствительны к голоданию (Austad and Fischer, 2016) и данные не показаны). Самки претерпевают метаболический сдвиг при спаривании, чтобы максимально увеличить кладку яиц, что изменяет их предпочтения в составе пищи и траекторию старения (Lee et al., 2008). Наконец, самки существенно отличаются от самцов по своим циркадным часам и двигательному поведению, особенно после спаривания (Fujii et al., 2007; Helfrich-Förster, 2000; Isaac et al., 2010). Будет интересно узнать, какие аспекты женской физиологии делают их более восприимчивыми к TRF. Наше открытие, что TRF, в принципе, работает и у мужчин, предполагает, что корректировка продолжительности TRF, фазы или состава пищи может создать условия для более универсальной реакции мужчин.
, 2019). Хотя мы не знаем, почему только спарившиеся самки показывают увеличение продолжительности жизни с TRF, различные соображения могут играть роль в дифференциальной реакции спарившихся самок.Самки мух крупнее самцов, демонстрируют различия в метаболизме и менее чувствительны к голоданию (Austad and Fischer, 2016) и данные не показаны). Самки претерпевают метаболический сдвиг при спаривании, чтобы максимально увеличить кладку яиц, что изменяет их предпочтения в составе пищи и траекторию старения (Lee et al., 2008). Наконец, самки существенно отличаются от самцов по своим циркадным часам и двигательному поведению, особенно после спаривания (Fujii et al., 2007; Helfrich-Förster, 2000; Isaac et al., 2010). Будет интересно узнать, какие аспекты женской физиологии делают их более восприимчивыми к TRF. Наше открытие, что TRF, в принципе, работает и у мужчин, предполагает, что корректировка продолжительности TRF, фазы или состава пищи может создать условия для более универсальной реакции мужчин.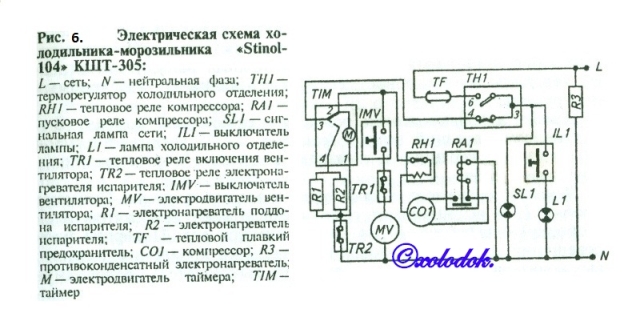
TRF, циркадные часы и режимы освещения
Мы наблюдаем, что TRF-опосредованное увеличение продолжительности жизни при определенных условиях аналогично DR (Katewa et al., 2015; Ulgherait et al., 2016) – зависят от неповрежденных циркадных часов, в то время как при определенных условиях освещения эффекты TRF могут быть независимыми от часов. В циклах LD часовые мутанты на 0 и tim 01 не демонстрируют увеличенной продолжительности жизни в ответ на TRF по сравнению с ALF и постоянного света, что вызывает аритмию у животных дикого типа. , также отменяет увеличение продолжительности жизни за счет TRF, предполагая зависимость от циркадных часов. Интересно, что постоянные условия – DD или LL – позволяют часовым мутантам получать пользу от TRF, что указывает на преимущества TRF, не зависящие от часов, как описано ранее (Chaix et al., 2019a; Xu et al., 2011) и свет как вторичный параметр. Свет: темные циклы представляют собой стимул, организующий суточные ритмы у часовых мутантов независимо от циркадных часов, и вызывают профиль экспрессии генов, отличный от, но частично перекрывающийся с профилем, управляемым циркадными часами (Wijnen et al. , 2006). С другой стороны, кормление по времени – это цейтгебер, который может увеличивать циркадные ритмы в метаболических тканях Drosophila с интактными часами или без них (Xu et al., 2011) и увеличивать амплитуду метаболических параметров и циклов сна / бодрствования в часах. меньше мышей (Chaix et al., 2019a), или, если они представлены в неестественное время, отрицательно влияют на другие физиологические параметры (Xu et al., 2011). Наши наблюдения могут быть объяснены моделью, в которой циркадные часы, свет и пищевые импульсы взаимодействуют сложным образом: программы экспрессии генов, запускаемые LD и LL, различаются у мутантов дикого типа и тактовых мутантов. Программы экспрессии генов, индуцированные LD у часовых мутантов и LL у мух дикого типа, противостоят полезным эффектам TRF, тогда как LD у дикого типа и DD во всех генотипах совпадают с эффектами TRF, способствующими долголетию.Для полного понимания этого взаимодействия необходимы дальнейшие исследования. Было показано, что сам свет играет важную роль в старении.
, 2006). С другой стороны, кормление по времени – это цейтгебер, который может увеличивать циркадные ритмы в метаболических тканях Drosophila с интактными часами или без них (Xu et al., 2011) и увеличивать амплитуду метаболических параметров и циклов сна / бодрствования в часах. меньше мышей (Chaix et al., 2019a), или, если они представлены в неестественное время, отрицательно влияют на другие физиологические параметры (Xu et al., 2011). Наши наблюдения могут быть объяснены моделью, в которой циркадные часы, свет и пищевые импульсы взаимодействуют сложным образом: программы экспрессии генов, запускаемые LD и LL, различаются у мутантов дикого типа и тактовых мутантов. Программы экспрессии генов, индуцированные LD у часовых мутантов и LL у мух дикого типа, противостоят полезным эффектам TRF, тогда как LD у дикого типа и DD во всех генотипах совпадают с эффектами TRF, способствующими долголетию.Для полного понимания этого взаимодействия необходимы дальнейшие исследования. Было показано, что сам свет играет важную роль в старении. Как режим освещения, так и цветовой состав света по-разному влияют на старение у Drosophila (Nash et al., 2019; Sheeba et al., 2000), поэтому будет интересно посмотреть, как индуцированные светом молекулярные пути вносят вклад в эффекты TRF. Хотя адаптивное значение циркадных часов было продемонстрировано у различных форм жизни, включая плодовых мушек и людей (Patke et al., 2019), роль часовых генов в долголетии более сложна. Удивительно, хотя мутанты продолжительности сна у Drosophila оказывают сильное влияние на продолжительность жизни, мутанты циркадных часов у Drosophila и мыши не обнаруживают значительного дефицита продолжительности жизни (Dubrovsky et al., 2010; Klarsfeld and Rouyer, 1998; Kondratov et al., 2006), а при определенных условиях даже продлевают продолжительность жизни (Ulgherait et al., 2020). Будущие исследования покажут, как продлевающие и сокращающие жизнь аспекты пищи, света и часов взаимодействуют на механистическом уровне, чтобы определить продолжительность жизни животного.
Как режим освещения, так и цветовой состав света по-разному влияют на старение у Drosophila (Nash et al., 2019; Sheeba et al., 2000), поэтому будет интересно посмотреть, как индуцированные светом молекулярные пути вносят вклад в эффекты TRF. Хотя адаптивное значение циркадных часов было продемонстрировано у различных форм жизни, включая плодовых мушек и людей (Patke et al., 2019), роль часовых генов в долголетии более сложна. Удивительно, хотя мутанты продолжительности сна у Drosophila оказывают сильное влияние на продолжительность жизни, мутанты циркадных часов у Drosophila и мыши не обнаруживают значительного дефицита продолжительности жизни (Dubrovsky et al., 2010; Klarsfeld and Rouyer, 1998; Kondratov et al., 2006), а при определенных условиях даже продлевают продолжительность жизни (Ulgherait et al., 2020). Будущие исследования покажут, как продлевающие и сокращающие жизнь аспекты пищи, света и часов взаимодействуют на механистическом уровне, чтобы определить продолжительность жизни животного.
Мы наблюдаем, что ген часов tim 01 не требуется в нейронах для эффектов долголетия TRF, что указывает на то, что периферических циркадных часов достаточно для эффектов TRF. Этот результат согласуется с предыдущими результатами на мышах, показывающими, что кормление по времени действует на периферийные часы (Manoogian and Panda, 2017). TRF увеличивает циркадную амплитуду различных метаболических факторов как в печени мышей, так и в жировом теле мух (печень мух), в то время как циркадные ритмы в головном мозге остаются неизменными (Xu et al., 2011). Кроме того, у мышей еда, не совпадающая по фазе с циклом сна / бодрствования, может создавать циркадный диссонанс в организме, при этом часы печени и мозга работают в разных фазах (Damiola et al., 2000). Будущие исследования будут направлены на определение органа (ов), в котором требуются функциональные часы, а также на определение молекулярных путей, обеспечивающих преимущества в отношении долголетия, посредством тканеспецифической функции часов.
Теории старения
Две теории старения постулируют, что старение – это либо накопление молекулярных повреждений после завершения репродукции – теория «одноразовой сомы» (Кирквуд, 1977), либо результат гиперпластических процессов – теория «Вздутая сома», или теория гиперфункции, согласно которой гиперплазия и гипертрофия приводят к патологиям в позднем возрасте (Gems and Partridge, 2013).TRF-опосредованное увеличение продолжительности жизни может влиять на старение через обе категории: циркадные часы, как было показано, увеличивают детоксифицирующий метаболизм ксенобиотиков и устойчивость к молекулярным повреждениям от активных форм кислорода (Gachon et al., 2006; Rakshit and Giebultowicz, 2013), предполагая, что TRF может улучшить старение за счет уменьшения накопления молекулярных повреждений. С другой стороны, наше исследование показывает, что TRF-мухи теряют вес, уменьшая биомассу и потенциально уменьшая гиперплазию и гипертрофию во время старения, подтверждая теорию вздутия сомы. Будущие исследования должны выявить точный молекулярный механизм TRF-опосредованного увеличения продолжительности жизни, потенциально создавая новые терапевтические мишени для борьбы со старением и возрастных патологий.
Будущие исследования должны выявить точный молекулярный механизм TRF-опосредованного увеличения продолжительности жизни, потенциально создавая новые терапевтические мишени для борьбы со старением и возрастных патологий.
Методы
Drosophila штаммов Isogenic Iso1CJ (Yin et al. 1994) использовали в качестве дикого типа. Циркадные мутанты на 0 и tim 01 (Konopka and Benzer, 1971; A. Sehgal et al., 1994) были использованы для изучения эффектов TRF на часовых мутантов. UAS-timRNAi и tub-Gal4 взяты из складского центра Блумингтона, № 29583 и № 5138, соответственно. nsyb-Gal4 – лабораторный инвентарь. Sleep мутанты inc1 (Stavropoulos and Young, 2011), fumin (Kume et al., 2005), sss P1 (Koh et al., 2008) и wakeD1 и wakeD2 (Liu et al., 2014) были использованы для анализа TRF на мутантах сна. Все мутанты были недавно (менее 2 лет) скрещены с мухами дикого типа в течение по крайней мере 5 поколений.
Анализ долголетия
Животных собирали после закрытия и давали возможность спариваться и стареть в течение 2 дней во флаконы, содержащие корм для мух, состоящий из 0,6% (мас. / Об.) Агара (Mooranger Inc # 41004), 5,25% (мас. / Об.) Желтого цвета. кукурузная мука (VWR # IC
180), 2,2% (масс. / об.) дрожжей (Lesaffre Yeast Co. # 73050), 5% (об. / об.) мелассы (Woolco Foods Inc. # 595650), 0,625% (об. / об.) тегосепт ( 30% гидробензойной кислоты (Sigma # H5501) в 95% этаноле), 0,66875% (об. / Об.) Пропионовой кислоты (VWR # JTU330-9). Затем животных слегка анестезировали CO 2 и разделяли по флаконам с животными того же пола, которые содержали в циклах LD при 25 ° C.Поскольку плотность популяции влияет на продолжительность жизни, а скученность укорачивает продолжительность жизни, мы поместили по 30 мух в каждую пробирку (Joshi and Mueller, 1997). Мух на TRF вручную переносили во флаконы с 1,1% агаром в конце легкой части цикла LD и возвращали обратно в стандартную кукурузную муку в начале легкой части цикла LD. Животных на ALF ежедневно переводили на свежий корм после того, как начальные эксперименты не показали разницы в продолжительности жизни между 12-часовым и 24-часовым переносом ALF (не показано). Ежедневно регистрировали количество погибших животных.Эксперименты в условиях постоянного освещения проводили в инкубаторе при 25 ° C и включении света на все время эксперимента. Эксперименты в постоянной темноте проводили в темном инкубаторе, и животных обрабатывали с красным светом в темной комнате для оценки выживаемости и перевода в новые флаконы. Мы использовали не менее трех технических повторов для каждого условия в каждом эксперименте, всего 90 мух на условие, что соответствует стандартам в данной области, например (Castillo-Quan et al., 2019). Маскирование использовалось во время сбора и анализа данных.Лог-ранговые тесты выживаемости (Mantel-Cox) выполняли в Graphpad Prism для сравнения выживаемости групп TRF и ALF.
Животных на ALF ежедневно переводили на свежий корм после того, как начальные эксперименты не показали разницы в продолжительности жизни между 12-часовым и 24-часовым переносом ALF (не показано). Ежедневно регистрировали количество погибших животных.Эксперименты в условиях постоянного освещения проводили в инкубаторе при 25 ° C и включении света на все время эксперимента. Эксперименты в постоянной темноте проводили в темном инкубаторе, и животных обрабатывали с красным светом в темной комнате для оценки выживаемости и перевода в новые флаконы. Мы использовали не менее трех технических повторов для каждого условия в каждом эксперименте, всего 90 мух на условие, что соответствует стандартам в данной области, например (Castillo-Quan et al., 2019). Маскирование использовалось во время сбора и анализа данных.Лог-ранговые тесты выживаемости (Mantel-Cox) выполняли в Graphpad Prism для сравнения выживаемости групп TRF и ALF.Анализ сна и циркадных ритмов
Животных соответствующего возраста из экспериментов TRF в LD загружали в стеклянные пробирки, содержащие стандартную кукурузную муку, и в течение четырех дней анализировали LD для измерения сна и в течение пяти дней в DD на ритмичность с использованием мониторов DAM5 (Trikinetics). Данные о двигательных способностях собирались в 1-минутные интервалы и 5-минутный период бездействия (Huber et al., 2004; Shaw et al., 2000) был определен как сон. Сон рассчитывался с использованием специального программного обеспечения на Python (Axelrod S. неопубликовано). Мертвые животные исключались из анализа либо с помощью программного обеспечения, либо путем визуального осмотра. Тесты Манна-Уитни были выполнены для сравнения сна между группами TRF и ALF с использованием Graphpad Prism.
Данные о двигательных способностях собирались в 1-минутные интервалы и 5-минутный период бездействия (Huber et al., 2004; Shaw et al., 2000) был определен как сон. Сон рассчитывался с использованием специального программного обеспечения на Python (Axelrod S. неопубликовано). Мертвые животные исключались из анализа либо с помощью программного обеспечения, либо путем визуального осмотра. Тесты Манна-Уитни были выполнены для сравнения сна между группами TRF и ALF с использованием Graphpad Prism.
Вес мухи
Для измерения веса мухи мы разрешили всем группам мух (ALF и TRF) оставаться на свежих кормах примерно на 10 часов. Мух разделяли на группы по 10 особей и взвешивали на весах (Mettler AT20).Приведены данные для трех или четырех экземпляров среднего веса 10 мух. Тесты Манна-Уитни были выполнены для сравнения веса между группами TRF и ALF с использованием Graphpad Prism.
Анализ кормления
Мы измерили кормление взрослых мух с помощью теста CApillary FEeder (CAFE) (Diegelmann et al. , 2017; Ja et al., 2007). 1-2-дневных животных отбирали и помещали в LD при 25 ° C. Курс 14- или 32-дневного TRF проводился в группах TRF, получавших пищу только от ZT 0-12 при включенном свете, по сравнению с ALF с постоянным кормом. в равных условиях.После 14 или 32 дней TRF 10 мух помещали во флакон, покрытый ватным тампоном и капиллярной трубкой с пищей, и каждое измерение проводили в трех экземплярах. Животных помещали в пластиковые флаконы с 1,1% агаром и жидкую пищу, содержащую 5% сахарозы, доставляли с помощью боросиликатных капиллярных трубок. Капилляр оценивали каждые три часа на предмет продолжительности смены корма, и корм был перезагружен. Для каждого эксперимента строили стандартную кривую, определяя изменение длины корма в капиллярах для разных объемов корма.В качестве контроля испарения измерения были представлены как разница между флаконами с мухами и пищей и флаконами с пищей и без мух.
, 2017; Ja et al., 2007). 1-2-дневных животных отбирали и помещали в LD при 25 ° C. Курс 14- или 32-дневного TRF проводился в группах TRF, получавших пищу только от ZT 0-12 при включенном свете, по сравнению с ALF с постоянным кормом. в равных условиях.После 14 или 32 дней TRF 10 мух помещали во флакон, покрытый ватным тампоном и капиллярной трубкой с пищей, и каждое измерение проводили в трех экземплярах. Животных помещали в пластиковые флаконы с 1,1% агаром и жидкую пищу, содержащую 5% сахарозы, доставляли с помощью боросиликатных капиллярных трубок. Капилляр оценивали каждые три часа на предмет продолжительности смены корма, и корм был перезагружен. Для каждого эксперимента строили стандартную кривую, определяя изменение длины корма в капиллярах для разных объемов корма.В качестве контроля испарения измерения были представлены как разница между флаконами с мухами и пищей и флаконами с пищей и без мух.
Финансирование
Эта работа была поддержана Calico Life Sciences LLC, NIH, номера наград NS053087 и GM136237 (M.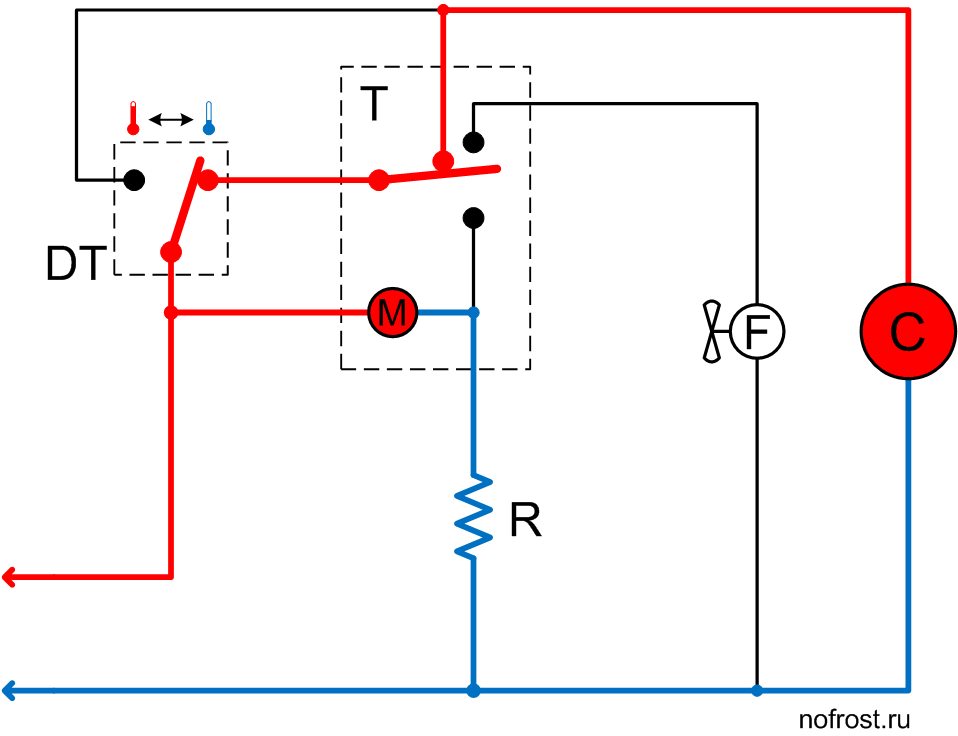 W.Y.), и грантом Программы обучения медицинских ученых (NIH), номер награды T32GM007739 (округ Колумбия).
W.Y.), и грантом Программы обучения медицинских ученых (NIH), номер награды T32GM007739 (округ Колумбия).
Вклад авторов
Проект был задуман S.A. и M.W.Y. S.A., D.C. и M.W.Y разработали эксперименты.D.C. проводил эксперименты под руководством S.A. и M.W.Y. и случайная помощь от S.A .. D.C. и S.A. проанализировала данные, которые обсуждались всеми авторами. S.A. написал рукопись при участии всех авторов.
Конкурирующие интересы
Авторы заявляют об отсутствии конкурирующих финансовых интересов.
Наличие данных и материалов
Представленные данные представлены в основном документе и дополнительных материалах.
Благодарности
Авторы благодарят С.Линкольну за помощь с TRF-переворотом и анализами долголетия, С. Джордану за полезные отзывы о рукописи, а также Р. Джексону, А. Сегалу, М. Ву и биржевому центру Блумингтона для запасов мух.
Принцип и применение таймера STM32
1 、 Классификация таймера
В серии stm32f1 есть 8 таймеров в дополнение к взаимосвязанным продуктам, которые делятся на базовый таймер, общий таймер и расширенный таймер. Базовые таймеры tim6 и tim7 – это 16-битные таймеры, которые могут только считать.Их можно синхронизировать только без внешнего io. Общий таймер TIM2 / 3/4/5 – это 16-битный таймер, который может вести обратный / обратный отсчет. Это может быть синхронизация, сравнение выходных данных и захват входных данных. Каждый таймер имеет четыре внешних входа. Расширенный таймер TIM1 / 8 – это 16-битный таймер, который может вести обратный / обратный отсчет. Это может быть синхронизация, сравнение выходов, захват входа, дополнительный выходной сигнал трехфазного двигателя, и каждый таймер имеет 8 внешних IO.
Базовые таймеры tim6 и tim7 – это 16-битные таймеры, которые могут только считать.Их можно синхронизировать только без внешнего io. Общий таймер TIM2 / 3/4/5 – это 16-битный таймер, который может вести обратный / обратный отсчет. Это может быть синхронизация, сравнение выходных данных и захват входных данных. Каждый таймер имеет четыре внешних входа. Расширенный таймер TIM1 / 8 – это 16-битный таймер, который может вести обратный / обратный отсчет. Это может быть синхронизация, сравнение выходов, захват входа, дополнительный выходной сигнал трехфазного двигателя, и каждый таймер имеет 8 внешних IO.
Ядром базового таймера является временная шкала, а также общий таймер и расширенный таймер.
1. Источник тактовой частоты
Тактовая частота таймера timxclk, то есть внутренние часы CK_ Int предоставляются после деления частоты предварительным делителем apb1. Если коэффициент предварительного делителя apb1 равен 1, частота остается неизменной. В противном случае, если частота умножается на 2, коэффициент предварительного делителя apb1 в библиотечной функции равен 2, то есть pclk1 = 36m, поэтому тактовая частота таймера timxclk = 36 * 2 = 72m.
2. Часы счетчика
После того, как часы таймера проходят через предварительный делитель PSC, то есть CK_ CNT, используемый для управления счетчиком.PSC – это 16-битный предварительный делитель, который может делить тактовый сигнал таймера timxclk на любое число от 1 до 65536.
Конкретный метод расчета: CK_ CNT = TIMxCLK / (PSC + 1)。
3. Счетчик
Счетчик CNT – это 16-битный счетчик, который может только подсчитывать, а максимальное значение счетчика составляет 65535. Когда счет достигает регистра автоматической перезагрузки, генерируется и очищается событие обновления, чтобы начать счет с нуля.
4. Регистр автоматической перезагрузки
Регистр автоматической перезагрузки arr – это 16-битный регистр, который содержит максимальное значение, которое может подсчитать счетчик.Когда счетчик достигает этого значения, если прерывание разрешено, таймер сгенерирует прерывание переполнения.
5. Расчет времени отсчета
Время таймера равно периоду прерывания счетчика, умноженному на количество прерываний. Счетчик в CK_ Управляемый CNT, время подсчета числа равно CK_. Обратное значение CLK равно: 1 / (timxclk / (PSC + 1)), а время генерации прерывания равно: 1 / (ck_clk * ARR). Если вы установите переменное время в программе обслуживания прерываний для записи количества прерываний, вы можете рассчитать, что необходимое нам время синхронизации равно: 1 / CK_ CLK * (ARR + 1) * время。
Счетчик в CK_ Управляемый CNT, время подсчета числа равно CK_. Обратное значение CLK равно: 1 / (timxclk / (PSC + 1)), а время генерации прерывания равно: 1 / (ck_clk * ARR). Если вы установите переменное время в программе обслуживания прерываний для записи количества прерываний, вы можете рассчитать, что необходимое нам время синхронизации равно: 1 / CK_ CLK * (ARR + 1) * время。
1.Определения
Установите время ожидания и выполните указанную аппаратную операцию по истечении времени ожидания.
Самая основная функция таймера – отсчет времени. Например, он может регулярно отправлять данные через последовательный порт, регулярно собирать рекламные данные или генерировать прямоугольный сигнал PWM. Таймер может генерировать ШИМ для управления состоянием двигателя, что является общей особенностью промышленного управления.
2. Микросхемы серии Stm32f4xx: 2 таймера расширенного управления (TIM1 и tim8), 10 общих таймеров (10tim2-tim5, tim9-tim14), 2 базовых таймера (tim6 и tim7) и 2 сторожевых таймера
Для получения подробной информации, пожалуйста, обратитесь к китайскому справочному руководству, чтобы понять его характеристики и принцип
3. Режим отсчета таймера:
Режим отсчета таймера:
Общий таймер может вести обратный отсчет, обратный отсчет и обратный отсчет в обоих направлениях.
① Режим обратного отсчета: счетчик ведет отсчет от 0 до значения автоматической загрузки (timx_arr), затем снова начинает отсчет с 0 и генерирует событие переполнения счетчика.
② Режим обратного отсчета: счетчик запускается с автоматически загруженного значения (timx_arr) и ведет обратный отсчет до 0, затем перезапускается с автоматически загруженного значения и генерирует событие переполнения счетчика.
③ Режим центрального выравнивания (счетчик вверх / вниз): счетчик начинает отсчет от 0 до автоматически загруженного значения – 1, генерирует событие переполнения счетчика, затем ведет обратный отсчет до 1 и генерирует событие переполнения счетчика; Затем снова начните отсчет с 0.
Как показано на рисунке:
4. Принцип работы таймера общего назначения:
● регистр счетчика (timx_cnt)
● регистр предварительного делителя (timx_psc)
● регистр автоматической перезагрузки (timx_arr)
В случае режима увеличения вверх, счетчик (CNT) добавляется к значению автоматической перегрузки (ARR) от 0, а предварительный делитель (PSC) уменьшается один раз.
Метод расчета времени:
Предположим, что частота составляет 42 м * 2 = = 84 м = = 84000000 = = 1 с / / 42 в зависимости от количества часов таймера, обычно это * 2
Таймер таймера 1 с генерирует прерывание:
ARR = 84000; // 1 мс
PSC = 1000; // 1000 * 1 мс == 1 с
Как показано на рисунке, тактовая частота основного таймера составляет 42 м, поэтому соответствующий * 2 равен 84 м = 8400000 = 1 с (arr * PSC)
Продвинутый 84 * 2 = 96м = 960000000 = 1с
——– ——–
5.Шаги реализации прерывания от таймера
// 5.1 таймер часов.
RCC_ APB1PeriphClockCmd ();
//5.2. Инициализируйте таймер и настройте arr и PSC. (структура декларации)
TIM_ TimeBaseInit ();
typedef struct
{
uint16_ t TIM_ Prescaler;
uint16_ t TIM_ CounterMode;
uint16_ t TIM_ Period;
uint16_ t TIM_ ClockDivision;
uint8_ t TIM_ RepetitionCounter;
} TIM_ TimeBaseInitTypeDef;
// 5. 3. Запускаем прерывание таймера, настраиваем NVIC (структуру декларации)
NVIC_ Init ();
Настроить прерывание NVIC
NVIC_ InitStructure.NVIC_ IRQChannel = TIM3_ IRQn; // Таймер 3 прерывания
NVIC_ InitStructure.NVIC_ IRQChannelPreemptionPriority = 0x01; // приоритет приоритетного обслуживания 1
NVIC_ InitStructure.NVIC_ IRQChannelSubPriority = 0x03; // Приоритет ответа 3
NVIC_ InitStructure.NVIC_ IRQChannelCmd = ENABLE;
NVIC_ Init (& NVIC_InitStructure); // ④ Инициализировать NVIC
TIM_ Cmd (TIM3 , ENABLE); // ⑤ Включить таймер 3
// 5.4 установка tim3_ Dier разрешить обновление прерывания
TIM_ ITConfig ();
// Таймер включения 5,5.
TIM_ Cmd ();
//5.6. Функция обслуживания прерывания записи.
TIMx_ IRQHandler ();
// Сервисная функция прерывания таймера 3
void TIM3_ IRQHandler (void)
{
Если (tim_getitstatus (tim3, tim_it_update) = = set) // прерывание переполнения
{
/ * Обработан сегмент программы * /
}
TIM_ ClearITPendingBit (TIM3 , TIM_IT_Update); // Сброс флага прерывания, бит
}
TIM3_ Int_ Init (5000-1,8400-1); // Тактовая частота таймера составляет 84 м, а коэффициент деления частоты равен 8400, поэтому частота счета 84 м / 8400 = 10 кГц составляет 500 мс для 5000 раз
void TIM3_ Int_ Init (u16 arr , u16 psc)
{
TIM_ TimeBaseInitTypeDef TIM_ TimeBaseInitStructure;
NVIC_ InitTypeDef NVIC_ InitStructure;
RCC_ APB1PeriphClockCmd (RCC_APB1Periph_TIM3 , ВКЛЮЧИТЬ); /// Включить часы tim3
TIM_ TimeBaseInitStructure. TIM_ Period = arr; // Автоматическая перезагрузка значения
TIM_ Period = arr; // Автоматическая перезагрузка значения
TIM_ TimeBaseInitStructure.TIM_ Prescaler = psc; // Таймер деления частоты
TIM_ TimeBaseInitStructure.TIM_ CounterMode = TIM_ CounterMode_ Up; // Режим подсчета
TIM_ TimeBaseInitStructure.TIM_ ClockDivision = TIM_ CKD_ DIV1;
TIM_ TimeBaseInit (TIM3 , & TIM_TimeBaseInitStructure); // Инициализировать tim3
TIM_ ITConfig (TIM3 , TIM_IT_Update , ENABLE); // Разрешить таймер 3 Обновить прерывание
TIM_ Cmd (TIM3 , ENABLE); // Включить таймер 3
NVIC_ InitStructure.NVIC_ IRQChannel = TIM3_ IRQn; // Таймер 3 прерывания
NVIC_ InitStructure.NVIC_ IRQChannelPreemptionPriority = 0x01; // приоритет приоритетного обслуживания 1
NVIC_ InitStructure.NVIC_ IRQChannelSubPriority = 0x03; // Подприоритет 3
NVIC_ InitStructure.NVIC_ IRQChannelCmd = ENABLE;
NVIC_ Init (& NVIC_InitStructure);
}
// Сервисная функция прерывания таймера 3
void TIM3_ IRQHandler (void)
{
Если (tim_getitstatus (tim3, tim_it_update) = = set) // прерывание переполнения
{
LED1 =! LED1; // переворот DS1
}
TIM_ ClearITPendingBit (TIM3 , TIM_IT_Update); // Сброс флага прерывания, бит
}
Подробное описание структуры инициализации таймера
В заголовочном файле стандартной библиотеки stm32f10x_ Tim. H устанавливает четыре структуры инициализации для периферийных устройств таймера, и базовый таймер использует только одну из них, Tim_ Timebaseinittypedef, остальные три объясняются в главе о расширенном таймере.
H устанавливает четыре структуры инициализации для периферийных устройств таймера, и базовый таймер использует только одну из них, Tim_ Timebaseinittypedef, остальные три объясняются в главе о расширенном таймере.
typedef struct {
uint16_ t TIM_ Prescaler; // Предделитель
uint16_ t TIM_ CounterMode; // Режим счета
uint32_ t TIM_ Period; // Цикл таймера
uint16_ t TIM_ ClockDivision; // деление часов
uint8_ t TIM_ RepetitionCounter; // Повторить калькулятор
} TIM_ TimeBaseInitTypeDef; один миллион двести тридцать четыре тысячи пятьсот шестьдесят семь
(1) TIM_ Prescaler: настройка предварительного делителя таймера.Источником синхронизации являются часы таймера через предделитель. Он устанавливает timx_ значение регистра PSC. Его можно установить от 0 до 65535, чтобы получить деление частоты от 1 до 65536.
(2) TIM_ Режим счетчика: режим счета таймера, который может быть прямым счетом, обратным счетом и тремя режимами выравнивания по центру. Базовый таймер может только считать, т.е. timx_ CNT может увеличиваться только с 0 без инициализации.
Базовый таймер может только считать, т.е. timx_ CNT может увеличиваться только с 0 без инициализации.
(3) TIM_ Period: цикл таймера, который на самом деле предназначен для установки значения регистра автоматической перезагрузки и обновления его в теневом регистре при генерации события.Устанавливаемый диапазон от 0 до 65535.
(4) TIM_ Clockdivision: деление тактовой частоты, установка тактовой частоты таймера CK_ Коэффициент деления частоты между внутренней частотой и тактовой частотой дискретизации цифрового фильтра. Базовый таймер не имеет этой функции и не требует настройки.
(5) TIM_ Счетчик повторений: счетчик повторений – это бит специального регистра расширенного регистра управления. Количество выходных ШИМ очень легко контролировать. Здесь нет настройки.
Хотя базовая структура инициализации таймера состоит из пяти элементов, вам нужно установить только два из них для базового таймера.
просмотров публикаций:
4
Принципы субъективного времени первого и второго порядка
PS65Ch37-Meck ARI 31 октября 2013 г. 17:48
17:48
Claassen DO, Jones CR, Yu M, Dirnberger G, Malone T., et al. 2013. Расшифровка влияния дисфункции мозжечка
и базальных ганглиев на точность и вариабельность моторного времени. Neuropsychologia 51: 267–74
Conners CK, Levin ED, Sparrow E, Hinton SC, Erhardt D, et al. 1996 г.Никотин и внимание у взрослых
Расстройство дефицита внимания с гиперактивностью (СДВГ). Psychopharmacol. Бык. 32: 67–73
Коуп Т.Е., Грубе М., Сингх Б., Бурх Д.Д., Гриффитс Т.Д. 2013. Базальные ганглии во времени восприятия: синхронизация
производительность при множественной системной атрофии и болезни Хантингтона. Neuropsychologia 83: e1
Cordes S, Meck WH. 2013. Порядковое суждение у крысы: понимание более длинных и более коротких для
сверхсекундных, но не субсекундных длительностей.J. Exp. Psychol. Генерал В прессе
Cordes S, Williams CL, Meck WH. 2007. Общие представления абстрактных величин. Curr. Прямой.
Psychol. Sci. 16: 156–61
Coull JT, Cheng RK, Meck WH. 2011. Нейроанатомические и нейрохимические основы времени.
2011. Нейроанатомические и нейрохимические основы времени.
Neuropsychopharmacology 36: 3–25
Coull JT, Hwang HJ, Leyton M, Dagher A. 2013. Дофаминергическая модуляция моторного времени у здоровых
добровольцев различается в зависимости от исходной доступности предшественника DA.Время Восприятие времени. 1: 77–98
Coull JT, Назарян Б., Видал Ф. 2008. Определение времени, хранение и сравнение длительности стимула задействуют дискретные
анатомических компонентов перцепционной временной сети. J. Cogn. Neurosci. 20: 2185–97
Coull J, Nobre A. 2008. Отделение явного хронирования от временного ожидания с помощью фМРТ. Curr. Opin.
Neurobiol. 18: 137–44
Коулл Дж. Т., Видаль Ф., Назарян Б., Макар Ф. 2003. Функциональная анатомия модуляции внимания времени
оценка.Science 303: 1506–8
Дали Ю., Земин Л. 1993. Многослойная нейронная сеть прямого распространения с адаптивным весом таблицы поиска. Схема.
Sys. IEEE 4: 2411–14
IEEE 4: 2411–14
Данкерт Дж., Фербер С., Пун С., Бродерик С., Стример С. и др. 2007. Заброшенное время: нарушение временного
восприятия мультисекундных интервалов при одностороннем игнорировании. J. Cogn. Neurosci. 19: 1706–20
Дидрихсен Дж., Иври Р. Б., Прессинг Дж. 2003. Вклад мозжечка и базальных ганглиев в интервальную синхронизацию.См.
Meck 2003, стр. 457–83
Droit-Volet S, Cl´
ement A, Wearden JH. 2001. Временная генерализация у детей от 3 до 8 лет. J. Exp.
Детская психол. 80: 271–88
Droit-Volet S, Fayolle S, Lamotte M, Gil S. 2013. Время, эмоции и воплощение времени. Время
Восприятие времени. 1: 99–126
Droit-Volet S, Meck WH. 2007. Как эмоции окрашивают наше восприятие времени. Trends Cogn. Sci. 11: 504–13
Дройт-Волет С., Мек У.Х., Пенни ТБ.2007. Сенсорная модальность и восприятие времени у детей и взрослых.
Behav. Процесс. 74: 244–50
Droit-Volet S, Wearden JH. 2001. Деление височной кости у детей. J. Exp. Детская психол. 80: 142–59
Деление височной кости у детей. J. Exp. Детская психол. 80: 142–59
Durstewitz D, Deco G. 2008. Вычислительная значимость переходной динамики в корковых сетях. Евро. J.
Neurosci. 27: 217–27
Фальтер К.М., Норейка В., Уэрден Дж. Х., Бейли А. Дж.. 2012. Более последовательный, но менее чувствительный: интервалы времени в
расстройствах аутистического спектра.Q. J. Exp. Psychol. 65: 2093–107
Forstmann BU, Anwander A, Sch¨
afer A, Neumann J, Brown S, et al. 2010. Кортико-полосатые связи предсказывают
контроля над скоростью и точностью принятия перцептивных решений. Proc. Natl. Акад. Sci. USA 107: 15916–20
Fortin C, Fairhurst S, Malapani C, Morin C, Towey J, Meck WH. 2009. Ожидание у людей в мультисекундном интервале
пиковое время с промежутками. Atten. Восприятие. Психофизика. 71: 789–802
Fortin C, Schweickert R, Gaudreault R, Viau-Quesnel C.2010. На время влияют запросы в памяти
поиска, но не переключение задач. J. Exp. Psychol. Гм. Восприятие. Выполнять. 36: 580–95
J. Exp. Psychol. Гм. Восприятие. Выполнять. 36: 580–95
Фрейсс П. 1984. Восприятие и оценка времени. Анну. Rev. Psychol. 35: 1–36
Фристоун DM, MacInnis MLM, Church RM. 2013. Скорость ответа больше зависит от времени, чем от
на случай непредвиденных обстоятельств. Время Восприятие времени. 1: 3–20
Гешайдер Г.А. 1997. Психофизика: Основы. Махва, Нью-Джерси: Эрлбаум
Гиббон Дж.1977 г. Скалярное ожидание и закон Вебера для определения времени животных. Psychol. Rev. 84: 279–325
Гиббон Дж. 1991. Истоки скалярной теории времени. Учиться. Мотив. 22: 3–38
Гиббон Дж. 1992. Повсеместность скалярной синхронизации с часами Пуассона. J. Math. Psychol. 35: 283–93
Гиббон Дж., Чёрч, РМ. 1984. Источники дисперсии в теории обработки информации времени. У животных
Познание, изд. Х. Л. Ройтблат, Т. Г. Бевер, HS Terrace, стр. 465–88. Хиллсдейл, Нью-Джерси: Эрлбаум
www.Annualreviews.org • Свойства внутренних часов 763
Annu. Rev. Psychol. 2014. 65: 743-771. Загружено с сайта www.annualreviews.org
Rev. Psychol. 2014. 65: 743-771. Загружено с сайта www.annualreviews.org
Университетским колледжем Лондона 03.07.14. Только для личного пользования.
Итальянский TIM выпустил третье предупреждение о прибылях в 2021 году на фоне битвы в совете директоров
- TIM снизил прогноз по основной прибыли внутреннего бизнеса на 2021 год
- Совет директоров потратит больше времени на предложение KKR, сообщает источник
- Vivendi рассматривает вопрос о перестановках в совете директоров, говорят источники
МИЛАН, 16 декабря (Рейтер) – Telecom Italia (TIM) (TLIT.MI) выпустил третье предупреждение о прибылях в этом году перед заседанием совета директоров в пятницу, которое, как ожидается, займет больше времени, прежде чем действовать по предложению о поглощении от американского фонда KKR (KKR.N).
TIM, шестая по величине телекоммуникационная группа в Европе, заявила поздно в среду, что снизила прогноз прибыли на 2021 год для своего внутреннего бизнеса из-за более низких, чем ожидалось, доходов от фиксированной связи от партнерства с DAZN по показу итальянского футбола.
Главный акционер Vivendi обвинил бывшего генерального директора TIM Луиджи Губитози в плохих финансовых показателях телефонной группы и может потребовать капитального ремонта, чтобы вытеснить его из совета директоров, сообщили Reuters в среду четыре источника на условиях анонимности.подробнее
Зарегистрируйтесь сейчас и получите БЕСПЛАТНЫЙ неограниченный доступ к Reuters.com
Зарегистрируйтесь
Правление соберется в пятницу, чтобы обсудить, среди прочего, предложение KKR о поглощении за 33 миллиарда евро (37,29 миллиарда долларов), но источник сказал, что это маловероятно произойдет любой крупный шаг вперед.
«Телеком не может сейчас оценить подход KKR, потому что для этого нет условий», – сказал источник, добавив, что решение предоставить фонду доступ к его бухгалтерским книгам было маловероятным.
Любое решение, возможно, придется отложить до выхода нового бизнес-плана группы в начале следующего года, сказал источник.
«Такой сценарий немного снижает вероятность заявки, проверяя терпение KKR», – отмечает аналитик Banca Akros Андреа Де Вита.
KKR, которая уже имеет долю в сети «последней мили» TIM FiberCop, запросила период должной осмотрительности, прежде чем формализовать предложение, поданное 19 ноября, но не установила крайнего срока. подробнее
Тупиковая ситуация с корпоративным управлением также может усложнить ситуацию.
Если Губитози не уйдет с поста директора на заседании в пятницу, Vivendi может потребовать созыва внеочередного собрания акционеров для назначения нового совета, сообщили два источника.
Обновление совета директоров также может быть вызвано отставкой большинства директоров.
акций TIM упали на 0,9 в четверг. Динамика акций
TIM по сравнению с индексом голубых фишек Италии TIM теперь ожидает «незначительного снижения» в 2021 году органической прибыли до вычета процентов, налогов, износа и амортизации после аренды (EBITDA-AL) для своего внутреннего бизнеса. В октябре прогнозируется “резкое однозначное снижение” для бизнеса.
В октябре прогнозируется “резкое однозначное снижение” для бизнеса.
«Этот новый прогноз предполагает очень отрицательный четвертый квартал (для основной прибыли внутреннего бизнеса) по сравнению с падением на 9,8% за девять месяцев, что приведет к потенциальному падению более чем на 20% в годовом исчислении», – Де Вита сказал.
DAZN DEAL
Ранее в этом году TIM заключила сделку на 1 миллиард евро со спортивным стриминговым сервисом DAZN, чтобы попытаться расширить свои услуги широкополосного доступа и платного ТВ. подробнее
TIM теперь стремится сократить стоимость сделки после того, как соглашение о распределении матчей высшей футбольной лиги Италии принесло меньше доходов, чем ожидалось.подробнее
Любые единовременные резервы будут определены в свете продолжающихся переговоров по соглашению DAZN и будут зарезервированы в 2021 году, говорится в заявлении TIM.
TIM заявила, что ожидает, что органический EBITDA-AL группы в 2021 году превысит 5,4 миллиарда евро благодаря росту ее бразильского бизнес-подразделения, который подтвержден на уровне “средне однозначных”.
EBITDA-AL составила 6,3 миллиарда евро в 2020 году, согласно годовому отчету TIM, и ранее группа сообщала о «средне-однозначном» падении в этом году.
Брокерская компания Jefferies сообщила, что консенсус по органической EBITDA-AL в 2021 году на уровне группы составил 5,8 млрд евро.
Ожидается, что после выплаты 435 миллионов евро за лицензии скорректированный консолидированный чистый финансовый долг компании составит около 17,6 миллиардов евро.
(1 доллар = 0,8849 евро)
Зарегистрируйтесь сейчас и получите БЕСПЛАТНЫЙ неограниченный доступ к Reuters.com
Зарегистрируйтесь
Дополнительная информация от Шивани Танна, Вишала Вивека в Бангалоре и Стивена Джукса в Милане Под редакцией Кирстен Донован, Пола Симао и Лесли Адлер
Наши стандарты: принципы доверия Thomson Reuters.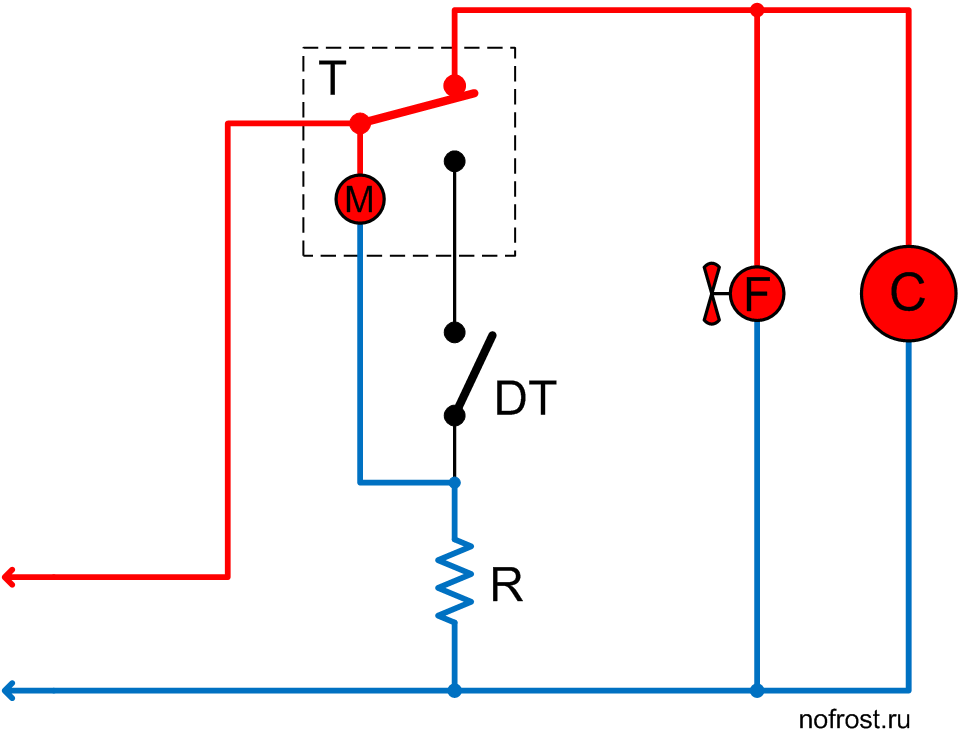
Базовый принцип и применение таймера STM32: обеспечивает точную задержку
Ранее мы говорили о системном тиковом таймере (systick). Вот таймер (ТИМ).
Различия между ними заключаются в следующем:
1. Systick – это периферийное устройство ядра. TIM – это периферийное устройство на кристалле.
2. Значение счетчика systick составляет 24 бита, и он может только обратный отсчет. Значение счетчика TIM составляет 16 бит, и есть счетчик с повышением (TIM6 / 7), который может увеличиваться или уменьшаться (другие счетчики, кроме TIM6 / 7).
3. Systick обычно обслуживает операционную систему.
В серии STM32F1, помимо взаимосвязанных продуктов, имеется всего 8 таймеров, разделенных на базовые, общие и расширенные.
Базовые таймеры TIM6 и TIM7 – это 16-битные таймеры, которые могут только подсчитывать и могут синхронизироваться только без внешнего ввода-вывода.
Таймер общего назначения TIM2 / 3/4/5 – это 16-битный таймер, который может вести обратный / обратный отсчет.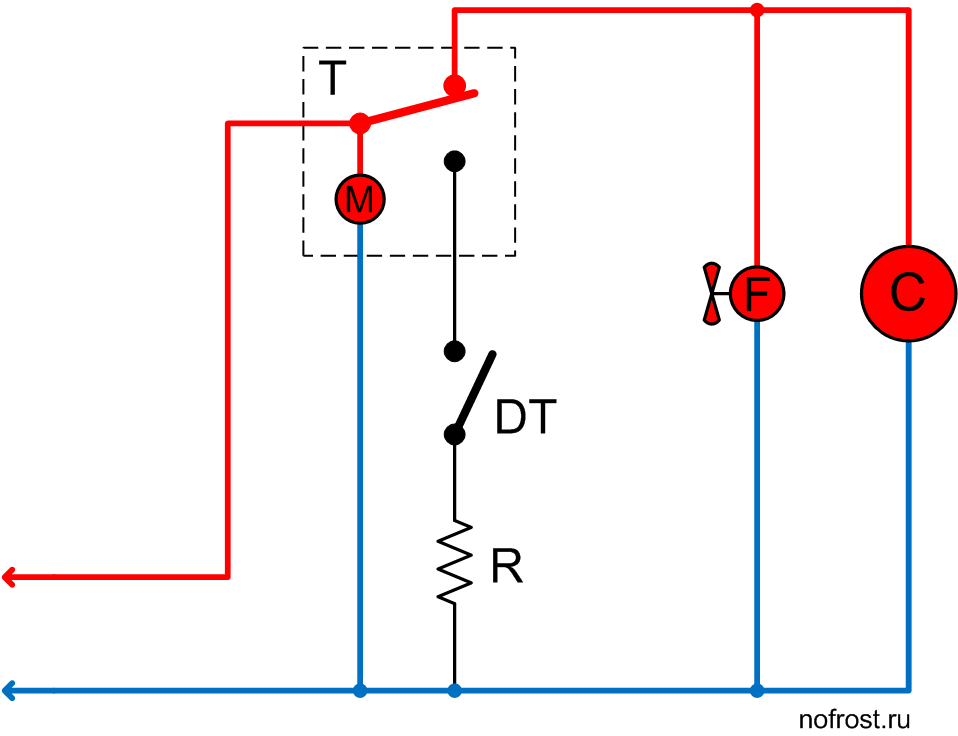 Это может быть синхронизация, сравнение выходных данных и захват входных данных. Каждый таймер имеет четыре внешних входа-выхода.
Это может быть синхронизация, сравнение выходных данных и захват входных данных. Каждый таймер имеет четыре внешних входа-выхода.
Расширенный таймер TIM1 / 8 – это 16-битный таймер, который может вести обратный / обратный отсчет. Это может быть синхронизация, сравнение выходов, захват входных сигналов и дополнительные выходные сигналы трехфазного двигателя. Каждый таймер имеет 8 внешних входов-выходов.
Базовые таймеры TIM6 и TIM7 могут также обеспечивать часы для ЦАП (цифро-аналоговый преобразователь).
объясняет внутреннюю структуру:
2.1 Источник тактовой частоты
Часы таймера TIMxCLK, которые являются внутренними часами CK_INT, делятся предварительным делителем APB1 и затем предоставляются.Если
Если коэффициент предварительного делителя APB1 равен 1, частота остается неизменной, в противном случае частота умножается на 2. Коэффициент предварительного делителя частоты APB1 в библиотечной функции равен 2, то есть PCLK1 = 36M, поэтому тактовая частота таймера TIMxCLK = 36 * 2 = 72М.
2.2 Счетчик часов
После того, как часы таймера проходят через предварительный делитель PSC, CK_CNT, он используется для управления счетчиком для подсчета. PSC – это 16-битный предварительный делитель, который может делить тактовый сигнал таймера TIMxCLK на любое число от 1 до 65536.
Конкретный метод расчета: CK_CNT = TIMxCLK / (PSC + 1)
2.3 Счетчик
Счетчик CNT – это 16-битный счетчик, который может только подсчитывать, а максимальное значение счета составляет 65535. Когда счет достигает регистра автоматической перезагрузки, генерируется событие обновления, и счетчик сбрасывается с начала.
2.4 Регистр автоматической перезагрузки
Регистр автоматической перезагрузки ARR – это 16-битный регистр, который содержит максимальное значение, которое может подсчитать счетчик.Когда счетчик достигает этого значения, если прерывание разрешено, таймер сгенерирует прерывание переполнения.
Вот одна точка, После переполнения таймера прерывание может быть сгенерировано или нет. Конкретная разница будет объяснена в следующей процедуре.
Конкретная разница будет объяснена в следующей процедуре.
Время одного отсчета таймера равно периоду прерывания счетчика, умноженному на количество прерываний.
Когда счетчик управляется CK_CNT, время для подсчета числа является обратным CK_CLK, которое равно: 1 / (TIMxCLK / (PSC + 1)), а время генерации прерывания равно: 1 / (CK_CLK * (ARR + 1)).
, т.е. (ARR + 1) (PSC + 1) / TIMxCLK。
Если в подпрограмме обработки прерываний установлено переменное время для записи количества прерываний, то мы можем вычислить, что необходимое нам время синхронизации равно:
(ARR + 1) (PSC + 1) / TIMxCLK * время。
Для периферийных устройств таймера установлены четыре структуры инициализации. Базовый таймер использует только один из них, а именно TIM_TimeBaseInitTypeDef. Остальные три объясняются в главе «Общий / расширенный таймер».
typedef struct {
uint16_t TIM_Prescaler;
uint16_t TIM_CounterMode;
uint32_t TIM_Period;
uint16_t TIM_ClockDivision;
uint8_t TIM_RepetitionCounter;
} TIM_TimeBaseInitTypeDef;
1. Источником тактовых импульсов предварительного делителя является тактовая частота таймера через прескалер, который устанавливает значение регистра TIMx_PSC. Регулируемый диапазон от 0 до 65535, достигая частотного деления от 1 до 65536.
2. Режим подсчета таймера, но это счет вверх, обратный отсчет и три режима с выравниванием по центру. Базовый таймер может только подсчитывать, то есть TIMx_CNT может увеличиваться только с 0 и не требует инициализации
3. Период таймера на самом деле предназначен для установки значения регистра автоматической перезагрузки, которое обновляется до Теневой регистр. .Диапазон настройки: от 0 до 65535.
4. Деление тактовой частоты, установите коэффициент деления частоты между тактовой частотой CK_INT таймера и тактовой частотой дискретизации цифрового фильтра. Базовые таймеры не имеют этой функции и не нуждаются в настройке.
5. Счетчик повторов – это специальный бит регистра расширенного регистра управления, который можно очень легко использовать для управления количеством выходных ШИМ. Не нужно здесь устанавливать.
Не нужно здесь устанавливать.
примечание: Теневой регистр Введение ARM – это особенность ARM.Сверху и под блок-схемой есть штриховка.
Это означает, что физически этот регистр соответствует двум регистрам: один – это регистр, который мы можем писать или читать, называемый регистром предварительной загрузки, а другой невидимый и не может быть Регистром, который на самом деле читает и записывает в него, но действительно работает в использование называется теневым регистром. В руководстве по данным описывается, что содержимое предварительно загруженного регистра может быть передано в теневой регистр в любое время, то есть два соединены (постоянно), или содержимое регистра предварительной загрузки передается в теневой регистр при каждом событии обновления. (UEV).
Как мы уже говорили, таймер может прерываться или не прерываться после переполнения таймера. Если он не прерывается, проверьте его флаг переполнения (TIM_FLAG_Update).
bsp_basetim. h содержание:
h содержание:
#ifndef __BSP_BASETIM_H
#define __BSP_BASETIM_H
#include "stm32f10x.h"
#define BASIC_TIM6
#ifdef BASIC_TIM6
#define BASIC_TIM TIM6
#define BASIC_TIM_APBxClock_FUN RCC_APB1PeriphClockCmd
#define BASIC_TIM_CLK RCC_APB1Periph_TIM6
#define BASIC_TIM_Period (1000-1)
#define BASIC_TIM_Prescaler (72-1)
#еще
#define BASIC_TIM TIM7
#define BASIC_TIM_APBxClock_FUN RCC_APB1PeriphClockCmd
#define BASIC_TIM_CLK RCC_APB1Periph_TIM7
#define BASIC_TIM_Period (1000-1)
#define BASIC_TIM_Prescaler 71
#endif
void BASIC_TIM_Config (недействительно);
void TIM_Delay_Ms (__IO uint32_t ms);
#endif
bsp_basetim.c содержание:
#include "bsp_basetim.h"
недействительным BASIC_TIM_Config (недействительным)
{
TIM_TimeBaseInitTypeDef TIM_TimeBaseStructure;
BASIC_TIM_APBxClock_FUN (BASIC_TIM_CLK, ВКЛЮЧИТЬ);
TIM_TimeBaseStructure.TIM_Period = BASIC_TIM_Period;
TIM_TimeBaseStructure.TIM_Prescaler = BASIC_TIM_Prescaler;
TIM_TimeBaseInit (BASIC_TIM, & TIM_TimeBaseStructure);
TIM_ClearFlag (BASIC_TIM, TIM_FLAG_Update);
TIM_Cmd (BASIC_TIM, ENABLE);
while (TIM_GetFlagStatus (BASIC_TIM, TIM_FLAG_Update) == RESET);
}
void TIM_Delay_Ms (__IO uint32_t мс)
{
uint32_t i;
для (я = 0; я <мс; я ++)
{
BASIC_TIM_Config ();
}
}
основной. c содержание:
c содержание:
#include "stm32f10x.h"
#include "bsp_led.h"
#include "bsp_basetim.h"
int main (пусто)
{
LED_GPIO_Config ();
в то время как (1)
{
LED_G (ВКЛ);
TIM_Delay_Ms (500);
LED_G (ВЫКЛ);
TIM_Delay_Ms (500);
}
}
1- Базовый таймер генерирует время 500 мс
2- Сделать инвертированный светодиод в функции прерывания
Содержимое bsp_basetim.h:
#ifndef __BSP_BASETIM_H
#define __BSP_BASETIM_H
#include "stm32f10x.h"
#define BASIC_TIM6
#ifdef BASIC_TIM6
#define BASIC_TIM TIM6
#define BASIC_TIM_APBxClock_FUN RCC_APB1PeriphClockCmd
#define BASIC_TIM_CLK RCC_APB1Periph_TIM6
#define BASIC_TIM_Period (1000-1)
#define BASIC_TIM_Prescaler (36000-1)
#define BASIC_TIM_IRQ TIM6_IRQn
#define BASIC_TIM_IRQHandler TIM6_IRQHandler
#еще
#define BASIC_TIM TIM7
#define BASIC_TIM_APBxClock_FUN RCC_APB1PeriphClockCmd
#define BASIC_TIM_CLK RCC_APB1Periph_TIM7
#define BASIC_TIM_Period (1000-1)
#define BASIC_TIM_Prescaler (36000-1)
#define BASIC_TIM_IRQ TIM7_IRQn
#define BASIC_TIM_IRQHandler TIM7_IRQHandler
#endif
недействительным BASIC_TIM_Init (недействительным);
#endif
Содержимое bsp_basetim. c:
c:
#include "bsp_basetim.h"
статическая пустота BASIC_TIM_NVIC_Config (void)
{
NVIC_InitTypeDef NVIC_InitStructure;
NVIC_PriorityGroupConfig (NVIC_PriorityGroup_0);
NVIC_InitStructure.NVIC_IRQChannel = BASIC_TIM_IRQ;
NVIC_InitStructure.NVIC_IRQChannelPreemptionPriority = 0;
NVIC_InitStructure.NVIC_IRQChannelSubPriority = 3;
NVIC_InitStructure.NVIC_IRQChannelCmd = ВКЛЮЧИТЬ;
NVIC_Init (& NVIC_InitStructure);
}
статическая пустота BASIC_TIM_Config (void)
{
TIM_TimeBaseInitTypeDef TIM_TimeBaseStructure;
BASIC_TIM_APBxClock_FUN (BASIC_TIM_CLK, ВКЛЮЧИТЬ);
TIM_TimeBaseStructure.TIM_Period = BASIC_TIM_Period;
TIM_TimeBaseStructure.TIM_Prescaler = BASIC_TIM_Prescaler;
TIM_TimeBaseInit (BASIC_TIM, & TIM_TimeBaseStructure);
TIM_ClearFlag (BASIC_TIM, TIM_FLAG_Update);
TIM_ITConfig (BASIC_TIM, TIM_IT_Update, ENABLE);
TIM_Cmd (BASIC_TIM, ENABLE);
}
недействительным BASIC_TIM_Init (недействительным)
{
BASIC_TIM_NVIC_Config ();
BASIC_TIM_Config ();
}
Содержание основной функции такое же.