Korf вентиляция отзывы: ООО “Корф” – отзывы о работе компании
Корф: отзывы сотрудников о работодателе
Почитал я тут отзывы и комментарии к отзывам… Что-то, видимо, зацепило, поэтому решил внести свою лепту. Вообще, плохая эта практика, нахваливать свою же фирму действующим работникам, тем более на руководящих должностях – это я в отношении последних двух комментов. Разит фальшью, пресные патриотические лозунги, “команда профессионалов”, “тщательный отбор и строгий конкурс” и тому подобное). Поэтому постараюсь быть объективным, к тому же много воды утекло с тех пор, как покинул Корф, и уже ничего с ним не связывает. Сразу отмечу, что работал я в инженерном отделе, поэтому вся моя публикация годится в основном для инженеров. За отдел продаж, клиентский отдел и другие – не скажу, там своя специфика, не вкурил, пусть другие пишут. Вот были высказывания, что офисы стремные. Ну да, не хоромы Брейвика, есть косяк, и вентиляция реально хреновая. С другой стороны, все необходимое есть, а если нет, предоставляется моментально, канцтовары не нужно выискивать по сусекам, сами периодически опрашивают, чего кому нужно.
 Приходилось мотаться в Дзержинский на маршрутках – втридорога и сплошь хамье за рулем.
Приходилось мотаться в Дзержинский на маршрутках – втридорога и сплошь хамье за рулем.Теперь по поводу самой работы. Да, придется вкалывать – ну а где не приходится? Глаза болят в конце дня? Ну да, болят, а у кого не болят? У бухгалтеров, проектировщиков, экономистов? На любом рабочем месте, где сидишь целый день и тупишь в монитор, главная рекомендация – время от времени переключать глаза на что-то другое. Кто-то еще упомянул, что перспектив нет… Ну блин, я вообще не знаю, откуда этот менталитет взялся, что все вокруг вам должны, и работодатели в том числе. На любом месте не будет перспектив, если грустно сидишь на попе и ждешь, когда все вокруг оглянутся на тебя и проникнутся, какой ты п…датый спец и вообще клевый парень (девушка). Я знаю людей, которые выросли внутри Корфа, потому что сами развивались, что-то предлагали, лезли во все дыры – их заметили. Всех замечают рано или поздно. Многие из тех, с кем я работал 3 года назад, работают там до сих пор, – вот вам и грандиозная текучка.

Работа в КОРФ ᐈ Отзывы сотрудников о работодателе КОРФ, зарплаты
Дате
Полезности
Рейтингу
Руководитель проекта
5,0 –
декабрь 2021
Что мне нравится в работодателе
Крупная производственная компания на рынке России Есть возможность и все условия для заработка без потолка. Выплачивают все бонусы,всё честно и прозрачно.
Что можно было бы улучшить
На мой взгляд, всё отлично.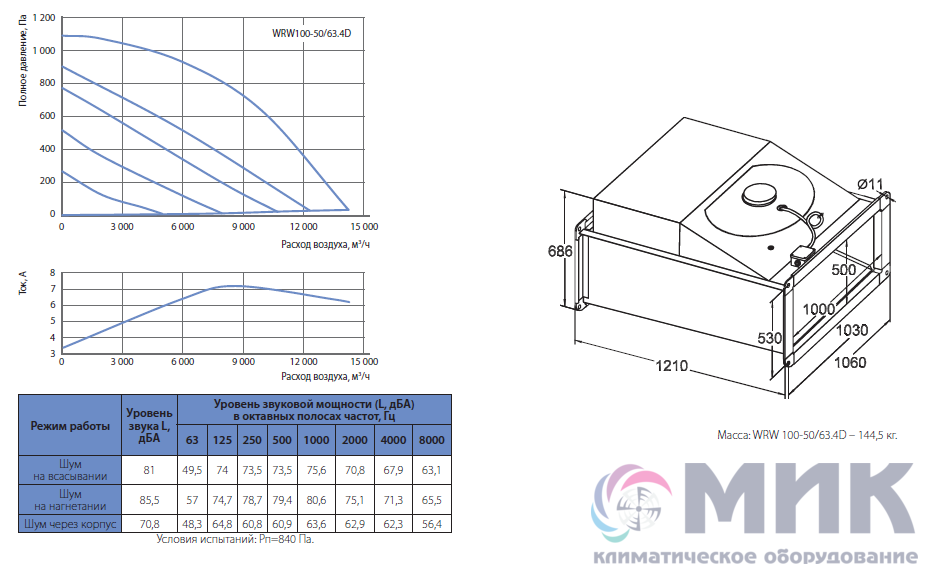 Руководство идёт на встречу! Поэтому компания не стоит на месте!!! ОП САМАРА/Волгоград.
Руководство идёт на встречу! Поэтому компания не стоит на месте!!! ОП САМАРА/Волгоград.
Условия труда
Уровень дохода
Условия для отдыха
Возможности ростаСвоевременная оплата труда
Удобное расположение работы
Удаленная работа
Оплата больничного
Компенсация транспортных расходов
Профессиональное обучение
Оплата мобильной связи
Система наставничества
Корпоративные мероприятия
РаскрытьСкрыть список льготЧасто задаваемые вопросы о работе в компании КОРФ
Рекомендуют ли сотрудники компанию КОРФ для трудоустройства?
Какая оплата труда в компании КОРФ
Сотрудники компании КОРФ оценивают оплату труда на 5 из 5, на основании 1 оценки.
Как сотрудники КОРФ оценивают начальство компании?
Сотрудники компании КОРФ оценивают работу начальства на 5 из 5, на основании 1 оценки.
Как сотрудники КОРФ оценивают карьерный рост в компании?
Сотрудники компании КОРФ оценивают карьерный рост в компании на 5 из 5, на основании 1 оценки.
Компания «Корф» основана в 2000 г. Основное направление деятельности- производство, разработка, изготовление оборудования для систем вентиляции и кондиционирования. Продукция КОРФ изготавливается на собственном производстве. Производственные мощности расположены в г. Дзержинский, Московской области.
Дзержинский, Московской области.
Сегодня КОРФ один из крупнейших производителей в России. Вся без исключения продукция КОРФ сертифицирована по ГОСТ, а также КОРФ сертифицирован по стандарту ISO 9001:2000.
Благодаря высокой технической оснащенности и налаженности производства, КОРФ производит широкий спектр канального оборудования и центральных кондиционеров. За 2005 г. нами было произведено более 14тыс различных наименований продукции. Оборудование КОРФ успешно конкурирует с известными брендами и является импортозамещающим. Производство КОРФ оснащено высокопроизводительным и высокоточным оборудованием с автоматизированным управлением TRUMF (Германия).
Коллектив Корфа насчитывает 200 человек сплоченных профессионалов любящих свое дело. Политика Корф: комплексно решать задачи и проблемы, с которыми сталкиваются наши клиенты. И по этому специалистам, работающим в Корф, уделяется особое внимание.
Все сотрудники постоянно проходят обучение и аттестацию на соответствие знаний и навыков занимаемой должности. Каждый сотрудник, начиная с производства и заканчивая отгрузкой, нацелен на оперативное обслуживание наших клиентов. Обращаясь в Корф Вам не придется беспокоится, обо всем побеспокоятся специалисты Корф.
Каждый сотрудник, начиная с производства и заканчивая отгрузкой, нацелен на оперативное обслуживание наших клиентов. Обращаясь в Корф Вам не придется беспокоится, обо всем побеспокоятся специалисты Корф.
Объекты Анкубера – Анкубера
Приглашаем к сотрудничеству
Для эффективного и взаимовыгодного сотрудничества мы готовы предложить:
- Конкурентные условия поставок оборудования Haier, Dantex, Marsa, Aerostar, Volcano, Flowair на территории ПФО
- Контроль уровня цен на оборудование марок Haier, Dantex, Marsa, Aerostar, Volcano, Flowair в регионе
- Отгрузка оборудования со склада в г.Самара
- Наличие бытовой линейки оборудования Haier, Dantex, Marsa, Aerostar, Volcano, Flowair на складе в Самаре
- Проектную поддержку перед производителем
- Помощь в продажах и маркетинге.


Стань дилером оборудования Haier, Dantex, Marsa, Aerostar, Volcano, Flowair!
Свяжитесь с нами и мы сможем предложить Вам выгодные условия сотрудничества!
Объекты Анкубера
Город: Самара
Оборудование: Volcano – воздушно-отопительные агрегаты. Вентиляционное оборудование Aerostar.
Город: Самара
Оборудование: Чиллеры Carrier, драйкулеры Nordvent.
Город: Самара
Оборудование: Volcano – воздушно-отопительные агрегаты.
Город: Самара
Оборудование: Чиллеры и фанкойлы Trane, общей холодильной мощностью 1300 кВт. Холодильный центр, приточно-вытяжные системы вентиляции.
Город: Самара
Оборудование: Volcano – воздушно-отопительные агрегаты.
Город: Самара
Оборудование: Мультизональная система кондиционирования Haier MRV II -C2: 2 внешних блока VRF суммарной производительностью 92 кВт и 33 внутренних блока VRF.
Город: Самара
Оборудование: Friterm – воздушный конденсатор системы холодильной централи торгового оборудования.
Город: Самара
Оборудование: Системы кондиционирования Haier. Вентиляционное оборудование Korf.
Город: Самара
Оборудование: Системы кондиционирования на базе руфтопов York, мультизональные системы Haier. Volcano – воздушно-отопительные агрегаты. Вентиляционное оборудование Korf.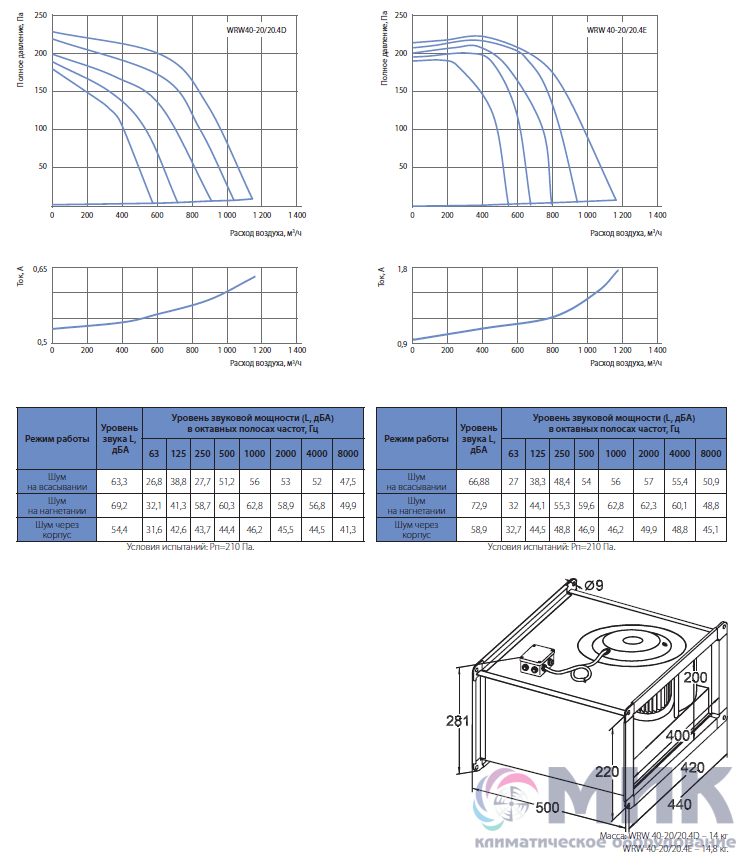
Город: Самара
Оборудование: Система промышленной очистки воздуха “СовПлим”
Город: Самара
Оборудование: Вытяжные системы Aerostar
Город: Самара
Оборудование: Flowair – воздушно-отопительные агрегаты.
Город: Самара
Оборудование: Haier – системы кондиционирования, Volcano – воздушно-отопительные агрегаты. Вентиляционное оборудование Korf.
Город: Бузулук
Оборудование: Haier – мультизональная система кондиционирования.
Город: Самара
Оборудование: Системы кондиционирования Haier. Вентиляционное оборудование Korf. Осушители Danterm. Тепловые завесы тропик.
Город: Самара
Оборудование: Мультизональныя системы кондиционирования VRF суммарной производительностью 160 кВт.
Город: Самара
Оборудование: Haier – бытовые системы кондиционирования и системы кондиционирования полупромышленного назначения. Приточно-вытяжные системы вентиляции.
Город: Самара
Оборудование: Мультизональная система кондиционирования Haier MRV 5. 2 внешних блока VRF суммарной производительностью 95 кВт и 29 внутренних блока VRF.
Город: Самара
Оборудование: VRF системы Haier общей холодопроизводительностью 260 кВт.
Город: Самара
Оборудование: На объекте смонтированы полупромышленные системы кондиционирования Haier общей холодопроизводительностью 896 кВт. Также специалистами нашей компании на этом объекте производился монтаж систем вентиляции, отопления, теплового пункта, освещения и автоматизации.
Город: Кинель-Черкассы
Оборудование: На объекте смонтированы полупромышленные системы кондиционирования Haier общей холодопроизводительностью 224 кВт.
Также специалистами нашей компании на этом объекте производился монтаж систем вентиляции, отопления, теплового пункта, освещения и автоматизации.
Город: Оренбург
Оборудование: На объекте смонтированы крышные кондиционеры Mitsubishi Electric r общей холодопроизводительностью 300 кВт. Также специалистами нашей компании на этом объекте производился монтаж систем вентиляции и автоматизации.
Город: Липецк
Оборудование: На объекте смонтированы чиллер, гидромодуль, охлаждающие ячейки (холодильная мощность 800 кВт), а также произведена реконструкция приточно-вытяжной вентиляции в цехе зерненого творога с монтажом приточно-вытяжной установки на 30 000 куб/час , тепло и холодоузлов, тканевых воздуховодов.
Город: Самара
Оборудование: VRF системы Haier общей холодопроизводительностью 182 кВт.
Город: Ульяновск
Оборудование: VRF системы Haier общей холодопроизводительностью 182 кВт.
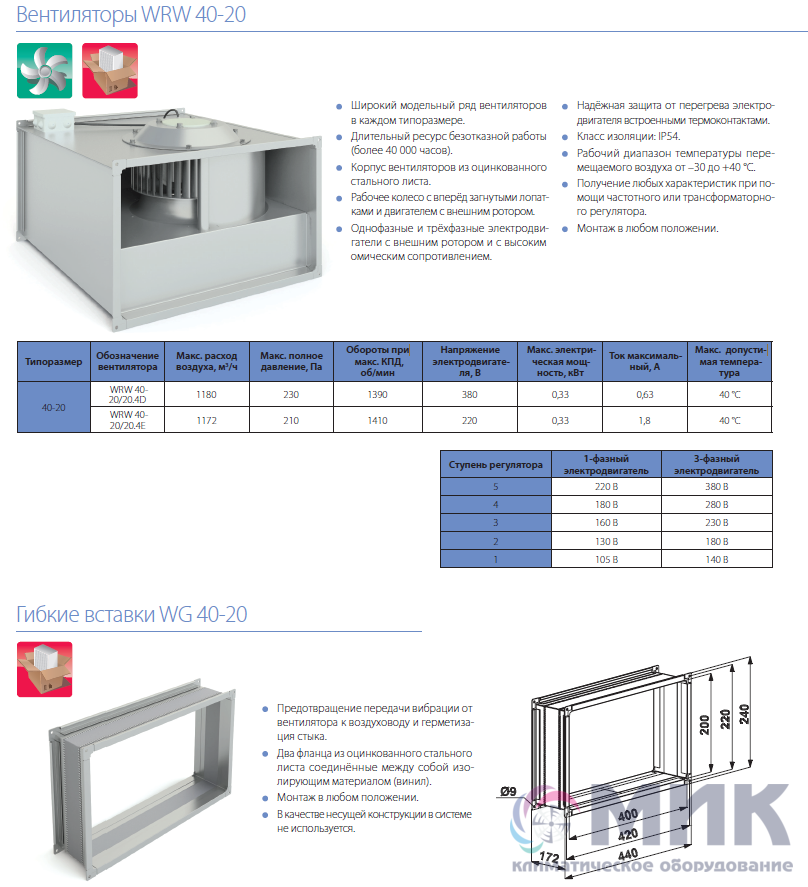
Приточные установки Korf
Ещё в совсем недалеком прошлом все влияние на климат в помещении ограничивалось вентиляторами в жаркое время и центральным отоплением в холодный сезон, а такие понятия, как кондиционер или UFO-обогреватель, широкому кругу потребителей были неизвестны. Научно-технический прогресс всего за каких-то десять лет изменил наш быт до неузнаваемости и превратил вчерашние неуютные квартиры в островки удобства и комфорта. Разумеется, коснулись инновации и вентиляционных систем, заменив обычные воздуховоды с решетками на высокотехнологичные устройства, которые не только забирают воздух с улицы, но и при необходимости нагревают зимой, охлаждают летом и фильтруют от пыли.
Одной из подобных систем стали вентиляционные приточные установки Korf. ПО «Корф» является крупным отечественным изготовителем центральных кондиционеров и канального оборудования в очень широком ассортименте, где высокое качество, исключительная функциональность и приемлемая цена обуславливают неизменную востребованность продукции и позволяют поддерживать репутацию Корф на должном уровне не только в условиях российского рынка, но и в Европе.
Вентиляционные приточные установки Корф представляют собой оптимальное решение для организации систем кондиционирования в зданиях самого разного назначения: от административных и офисных до промышленных. Диапазон производительности приточных систем Korf весьма широк, начиная от нескольких сотен и вплоть до тысяч кубометров воздуха в час. Это дает возможность обслуживать как отдельные комнаты жилых домов, так и помещения большей площади. Благодаря продуманной конструкции их монтаж может быть выполнен различными способами, что расширяет сферу их применения.
Что касается конструкционных особенностей, то вентиляционные приточные установки Korf могут быть сборного или моноблочного типа, где монтаж первого типа осуществляется по принципу конструктора и отличается сравнительно невысокой стоимостью, тогда как второй вариант более удобен при монтаже, компактен, обладает более низким уровнем шума при работе, но, соответственно, более высокой ценой.
Вентиляционные приточные установки Корф имеют следующий принцип действия:
– При поступлении воздуха он проходит через систему фильтров, где предварительно очищается.
– Уже очищенный воздух подается в помещение и охлаждается (летний вариант), или подаче предшествует предварительное нагревание (зимний вариант).
– Регулирования скорости и объема воздушных масс осуществляется за счет применения специальных клапанов, а уровень шума снижается при помощи установки шумопоглотителей, рассчитанных на поток воздуха до 20 м/с. Не так давно в ассортименте продукции появились принципиально новые звукоизолированные модели воздуховодов.
Не так давно в ассортименте продукции появились принципиально новые звукоизолированные модели воздуховодов.
Вне зависимости от выбранной модели, все вентиляционные приточные установки Korf обладают следующими достоинствами и преимуществами:
– В помещение поступает гарантированно свежий воздух, что благотворным образом сказывается на здоровье людей, нормализует влажность, снижает уровень пыли, способствует удалению неприятных запахов или вредных примесей.
– Системы Korf очень просты в работе. Достаточно настроить и запустить установку один раз, и ее бесперебойное функционирование обеспечено на длительный срок.
– Благодаря сравнительной простоте конструкции и отсутствию энергозатратных механизмов вентиляционные приточные установки Корф являются достаточно экономичными.
– Возможность монтажа с учетом всех архитектурных особенностей помещения, в том числе разработка индивидуальных проектов. Так, в каждом конкретном случае комплектация может зависеть от текущих задач и планируемого бюджета.
Так, в каждом конкретном случае комплектация может зависеть от текущих задач и планируемого бюджета.
Таким образом, вентиляционные приточные установки Korf на данный момент представляют собой оборудование, приобретение которого выгодно с позиции как цены, так и функциональности, обеспечивающей длительный срок работы без необходимости ремонта и замены комплектующих.
Калорифер korf, цена , водонагреватели промышленные от “ТД КОМТЕХ”
Торговый дом “Комтех” осуществляет поставку всего спектра промышленного тепловентиляционного оборудования.
В ассортименте более 1000 наименований: осевые и радиальные вентиляторы, тягодутьевые машины, крышные вентиляторы, водяные, паровые и электрические калориферы, воздушно-отопительные агрегаты, тепловые завесы и комплектующие к ним.
Вся продукция находится в наличии на нашем складе и может быть доставлена в любой регион России в кратчайшие сроки.
Мы гарантируем высокое качество наших изделий, подтвержденное сертификатами соответствия, и выгодные цены, как для оптовых, так и для розничных покупателей. Возможна отсрочка платежа!
Для получения консультации специалиста звоните по телефону +7 (343) 213-08-50
Вентиляционное оборудование:
Теплообменное оборудование:
Вам требуется поставка надежного теплообменного или вентиляционного промышленного оборудования? Ищете выгодные условия? Хотите получить заказ точно в срок?
Тогда звоните по телефону +7 (343) 213-08-50. Будем рады помочь Вам!
Наши преимущества
- Наличие более 1000 видов вентиляционного и теплового оборудования на складе Екатеринбурга.
- Выгодные цены, сравнимые с ценами конкурентов.
- Доставка по всем регионам Российской Федерации.
- Рассмотрение заявок в короткие сроки и своевременная доставка заказа.
- Дополнительные скидки на покупки оптом, в зависимости от объема товара, и отсрочку платежа.
- Качество товара гарантировано, предоставляется вся необходимая документация.

Товар в наличии на складе | Высокое качество | Оперативная доставка | Скидки оптовикам |
Схема работы
| Вы отправляете заявку | |
| Мы выставляем Вам счет | |
| Вы оплачиваете покупку удобным для Вас способом | |
| Получаете свой товар |
Вентиляция КОРФ
Вентиляция КОРФ представлена здесь наиболее популярными категориями оборудования, также Вы можете перейти в полный каталог вентиляции завода КОРФ или скачать прайс-лист.Вентиляционное оборудование московского завода KORF (КОРФ) является на сегодняшний день, пожалуй, самым современным и надёжным оборудованием на рынке вентиляционных технологий и систем кондиционирования. А начиналась деятельность этого завода с небольшого производства, организованного группой единомышленников, в арендованном боксе пожарной части в одном из подмосковных городов. Сейчас, по прошествии нескольких лет, производство КОРФ представляет собой высокотехнологичные процессы, практически полностью автоматизированные и использующие в своей работе новейшее немецкое оборудование. Добиться этого удалось благодаря дружной команде высококлассных специалистов, нацеленных на достижение единой цели, вывести на российский рынок вентиляционного оборудования высококачественный продукт, не уступающий по своим параметрам именитым европейским брендам. И это им удалось! Любой специалист, так или иначе связанный с рынком вентиляции, может подтвердить, что вентиляция КОРФ – это европейское качество, которое доступно в России.
А начиналась деятельность этого завода с небольшого производства, организованного группой единомышленников, в арендованном боксе пожарной части в одном из подмосковных городов. Сейчас, по прошествии нескольких лет, производство КОРФ представляет собой высокотехнологичные процессы, практически полностью автоматизированные и использующие в своей работе новейшее немецкое оборудование. Добиться этого удалось благодаря дружной команде высококлассных специалистов, нацеленных на достижение единой цели, вывести на российский рынок вентиляционного оборудования высококачественный продукт, не уступающий по своим параметрам именитым европейским брендам. И это им удалось! Любой специалист, так или иначе связанный с рынком вентиляции, может подтвердить, что вентиляция КОРФ – это европейское качество, которое доступно в России.
Чем же вентиляция КОРФ смогла привлечь к себе российского потребителя?
- Самое современное высокоточное производство
- Профессионалы на каждом этапе производства
- Европейское качество производимого оборудования
- Использование проверенных европейских комплектующих
- Самый полный ассортимент оборудования
- Индивидуальный подход к клиентам
- Высокий уровень качества сервиса
Грамотное сочетание вышеперечисленных качеств и позволило вывести продукцию КОРФ на первое место среди компаний, занимающихся подобной деятельностью. Наша компания “Линия Холода” уже достаточно давно занимается продажей, монтажом и сервисным обслуживанием холодильного оборудования для магазинов и очень часто наши клиенты спрашивали нас о возможности поставках вентиляционного оборудования, поскольку эти две сферы являются смежными. Мы долго думали, какого производителя вентиляции выбрать для представления в нашем интернет магазине холодильного оборудования, сравнивали цены, качество и сроки поставок оборудования, изучали отзывы специалистов и пришли к выводу, что на российском рынке, конкурентов у вентиляции КОРФ на данный момент просто нет.
Наша компания “Линия Холода” уже достаточно давно занимается продажей, монтажом и сервисным обслуживанием холодильного оборудования для магазинов и очень часто наши клиенты спрашивали нас о возможности поставках вентиляционного оборудования, поскольку эти две сферы являются смежными. Мы долго думали, какого производителя вентиляции выбрать для представления в нашем интернет магазине холодильного оборудования, сравнивали цены, качество и сроки поставок оборудования, изучали отзывы специалистов и пришли к выводу, что на российском рынке, конкурентов у вентиляции КОРФ на данный момент просто нет.
Итак, уважаемые посетители, рады Вам представить качественный продукт европейского уровня по российским ценам. Линейка продукции КОРФ имеет весьма большое количество позиций, если вы не нашли нужного оборудования на нашем сайте, позвоните нам по телефонам:
8 (495) 642 57 04
8 (926) 403 93 77
или напишите в форме связи, мы обязательно постараемся Вам помочь. И, конечно, же, у нас вы можете получить любую консультацию относительно холодильного оборудования для магазинов, а также систем бытового кондиционирования.
И, конечно, же, у нас вы можете получить любую консультацию относительно холодильного оборудования для магазинов, а также систем бытового кондиционирования.
VERTRO — российский производитель вентиляционного оборудования
Подбирает все основные типы климатического оборудования: для подбора доступно 7 категорий и 24 типа оборудования;
Современный и понятный интерфейс, в котором приятно работать;
Оперативный подбор: стандартная приточная система – всего за 30 секунд;
37 шаблонов самых ходовых установок для быстрого старта;
Подробные технические данные: более 70 параметров для установки с нагревателем и фильтром;
Оперативная поддержка технических специалистов и личного менеджера;
Быстрая подготовка коммерческого предложения для подобранного оборудования;
Доступна онлайн 24/7 из любой точки мира;
Более 400 пользователей-проектировщиков.
Перейти в программу
KORF Hydraulics — гибкие расчеты расхода жидкости
KORF Гидравлика – Гибкая жидкость расход расчеты 1. Резюме Korf Hydraulics — это мощная графическая программа для расчета расхода расхода и профилей давления в трубах и трубопроводных сетях.Гидравлика Korf отличается от других подобных программ тем, что пользователь может указать любую допустимую комбинацию расхода и давления, а также позволить Korf определить неизвестные величины. Единственное требование состоит в том, что единственное решение должно быть теоретически возможным. Korf может решить проблемы жидкости потока, начиная от одной трубы и заканчивая сложной двухфазной сетью труб, насосов, клапанов и других фитингов. 2. Принцип действия В литературе описано несколько методов расчета трубопроводных сетей, включая метод Харди-Кросса, Ньютона-Рафсона и метод линейной теории [1]. Хотя Корф был разработан независимо, он имеет некоторые общие характеристики с методом линейной теории. Метод Корфа быстр и почти всегда сходится с допустимыми спецификациями. Заметным исключением является контур, состоящий из насоса (или компрессора) с неизвестным расходом и напором, определяемым кривой насоса (или кривой компрессора).Только в этом случае Корф использует итерационный метод Эйткена-Стеффенсена для облегчения сходимости. Корф рассматривает все расходы и давление на входе и выходе всего оборудования как переменные или неизвестные. Чтобы определить эти неизвестные, Корф выполняет массовый баланс для каждой единицы оборудования и вычисляет падение давления для каждой трубы. Это так называемые внутренние спецификации. Даже с этими внутренними спецификациями уникальное решение проблемы невозможно без указания пользователем дополнительного числа расходов, давлений, перепадов давления, коэффициентов бета, Cv клапана и т.
2. Принцип действия В литературе описано несколько методов расчета трубопроводных сетей, включая метод Харди-Кросса, Ньютона-Рафсона и метод линейной теории [1]. Хотя Корф был разработан независимо, он имеет некоторые общие характеристики с методом линейной теории. Метод Корфа быстр и почти всегда сходится с допустимыми спецификациями. Заметным исключением является контур, состоящий из насоса (или компрессора) с неизвестным расходом и напором, определяемым кривой насоса (или кривой компрессора).Только в этом случае Корф использует итерационный метод Эйткена-Стеффенсена для облегчения сходимости. Корф рассматривает все расходы и давление на входе и выходе всего оборудования как переменные или неизвестные. Чтобы определить эти неизвестные, Корф выполняет массовый баланс для каждой единицы оборудования и вычисляет падение давления для каждой трубы. Это так называемые внутренние спецификации. Даже с этими внутренними спецификациями уникальное решение проблемы невозможно без указания пользователем дополнительного числа расходов, давлений, перепадов давления, коэффициентов бета, Cv клапана и т. д.Уникальное решение возможно, когда количество независимых спецификаций (включая внутренние спецификации) равно количеству переменных, как определено выше. Отслеживание этих спецификаций — нетривиальная задача для пользователя, но в Korf есть четыре функции, значительно упрощающие этот процесс. Во-первых, Korf постоянно подсчитывает и отображает количество спецификаций и переменных. Во-вторых, когда рисуется новая схема, Korf принимает определенные значения по умолчанию, что приводит к тому, что количество спецификаций всегда равно количеству переменных.Если пользователь впоследствии удалит один, новый должен быть добавлен где-то еще. В-третьих, Korf может отображать все текущие спецификации на чертеже под трубами/оборудованием. В-четвертых, при запуске задачи Korf C.J.C Kruger, KORF Hydraulics, сначала оценивает спецификации, чтобы определить, являются ли они независимыми. 3. Обсуждение и примеры. Верхняя схема на рисунке 1, которая является снимком экрана от Korf, представляет собой одну трубу.
д.Уникальное решение возможно, когда количество независимых спецификаций (включая внутренние спецификации) равно количеству переменных, как определено выше. Отслеживание этих спецификаций — нетривиальная задача для пользователя, но в Korf есть четыре функции, значительно упрощающие этот процесс. Во-первых, Korf постоянно подсчитывает и отображает количество спецификаций и переменных. Во-вторых, когда рисуется новая схема, Korf принимает определенные значения по умолчанию, что приводит к тому, что количество спецификаций всегда равно количеству переменных.Если пользователь впоследствии удалит один, новый должен быть добавлен где-то еще. В-третьих, Korf может отображать все текущие спецификации на чертеже под трубами/оборудованием. В-четвертых, при запуске задачи Korf C.J.C Kruger, KORF Hydraulics, сначала оценивает спецификации, чтобы определить, являются ли они независимыми. 3. Обсуждение и примеры. Верхняя схема на рисунке 1, которая является снимком экрана от Korf, представляет собой одну трубу. Корф проанализирует эту схему, чтобы иметь три переменные, а именно.давление подачи F1, давление продукта T1 и расход трубы L1. Он найдет только одну внутреннюю спецификацию, а именно. Падение давления в трубе L1. Таким образом, пользователь может указать любые две из перечисленных выше переменных, чтобы обеспечить уникальное решение. То есть давление на входе и выходе, или давление на входе и поток, или давление на выходе и поток. Для этого контура размер трубы следует оставлять неизвестным только в том случае, если указан расход, так как указание обоих давлений существенно фиксирует dP/длину и скорость, а изменение диаметра трубы не оказывает на них никакого влияния (Корф итерации до максимального или минимального размера трубы).Нижний контур на рисунке 1 представляет собой последовательный контур с тремя трубами, насосом и регулирующим клапаном. Корф будет анализировать эту схему, чтобы иметь девять переменных, а именно.
Корф проанализирует эту схему, чтобы иметь три переменные, а именно.давление подачи F1, давление продукта T1 и расход трубы L1. Он найдет только одну внутреннюю спецификацию, а именно. Падение давления в трубе L1. Таким образом, пользователь может указать любые две из перечисленных выше переменных, чтобы обеспечить уникальное решение. То есть давление на входе и выходе, или давление на входе и поток, или давление на выходе и поток. Для этого контура размер трубы следует оставлять неизвестным только в том случае, если указан расход, так как указание обоих давлений существенно фиксирует dP/длину и скорость, а изменение диаметра трубы не оказывает на них никакого влияния (Корф итерации до максимального или минимального размера трубы).Нижний контур на рисунке 1 представляет собой последовательный контур с тремя трубами, насосом и регулирующим клапаном. Корф будет анализировать эту схему, чтобы иметь девять переменных, а именно. давление подачи F2, давление продукта T2, давление на входе и выходе насоса P1, давление на входе и выходе клапана V1 и поток в трубах L2, L3 и L4. Корф найдет пять внутренних спецификаций, а именно. перепады давления в трубах L2, L3 и L4 и массовый баланс на насосе P1 и клапане V1. Таким образом, пользователь может указать любые четыре из перечисленных выше переменных, если они независимы.Независимость означает, что пользователь не может указать два массовых расхода расхода (что, очевидно, одинаково для последовательных труб), поскольку это эквивалентно балансу масс, который Korf выполняет внутри. Это также подразумевает, что по крайней мере одна спецификация должна быть давлением, поскольку Корфу нужно по крайней мере одно давление где-то в контуре, чтобы основывать на нем другие давления. В дополнение к перечисленным выше переменным, пользователь также может задать напор насоса прямо или косвенно через характеристику насоса.Аналогично для регулирующего клапана, где пользователь также может указать падение давления прямо или косвенно через клапан Cv.
давление подачи F2, давление продукта T2, давление на входе и выходе насоса P1, давление на входе и выходе клапана V1 и поток в трубах L2, L3 и L4. Корф найдет пять внутренних спецификаций, а именно. перепады давления в трубах L2, L3 и L4 и массовый баланс на насосе P1 и клапане V1. Таким образом, пользователь может указать любые четыре из перечисленных выше переменных, если они независимы.Независимость означает, что пользователь не может указать два массовых расхода расхода (что, очевидно, одинаково для последовательных труб), поскольку это эквивалентно балансу масс, который Korf выполняет внутри. Это также подразумевает, что по крайней мере одна спецификация должна быть давлением, поскольку Корфу нужно по крайней мере одно давление где-то в контуре, чтобы основывать на нем другие давления. В дополнение к перечисленным выше переменным, пользователь также может задать напор насоса прямо или косвенно через характеристику насоса.Аналогично для регулирующего клапана, где пользователь также может указать падение давления прямо или косвенно через клапан Cv. Кроме того, любой или все диаметры трубы могут быть неизвестны, если пользователь хочет, чтобы Korf определил ее размер на основе любого из пяти выбираемых критериев. Принципы этих примеров можно распространить на схемы практически с любой комбинацией труб, соединенных последовательно и параллельно. Две схемы, показанные на рис. 1, также могут относиться к одной и той же задаче, поскольку Корф по-прежнему сможет определить, имеет ли каждая из них уникальное решение.Таким образом, если вы преувеличиваете значение одного контура и занижаете значение другого контура, количество CACHE Page 1
Кроме того, любой или все диаметры трубы могут быть неизвестны, если пользователь хочет, чтобы Korf определил ее размер на основе любого из пяти выбираемых критериев. Принципы этих примеров можно распространить на схемы практически с любой комбинацией труб, соединенных последовательно и параллельно. Две схемы, показанные на рис. 1, также могут относиться к одной и той же задаче, поскольку Корф по-прежнему сможет определить, имеет ли каждая из них уникальное решение.Таким образом, если вы преувеличиваете значение одного контура и занижаете значение другого контура, количество CACHE Page 1
Влияние высоких доз витамина D на продолжительность искусственной вентиляции легких у пациентов в отделении интенсивной терапии
Иран Джей Фарм Рез. 2019 Весна; 18(2): 1067–1072.
МирМохаммад Мири
a Отделение интенсивной терапии, Медицинский и образовательный центр Эмама Хоссейна, Университет медицинских наук им. Шахида Бехешти, Тегеран, Иран.
Мехран Кучек
a Отделение интенсивной терапии, Медицинский и образовательный центр Эмама Хоссейна, Университет медицинских наук им. Шахида Бехешти, Тегеран, Иран.
Алиреза Рахат Дамарде
a Отделение интенсивной терапии, Медицинский и образовательный центр Эмама Хоссейна, Университет медицинских наук им. Шахида Бехешти, Тегеран, Иран.
Мохаммад Систанизад
b Кафедра клинической фармации, Факультет фармации, Университет медицинских наук им. Шахида Бехешти, Тегеран, Иран.
c Отделение фармацевтической помощи, Медицинский и образовательный центр Эмама Хоссейна, Университет медицинских наук им. Шахида Бехешти, Тегеран, Иран.
a Отделение интенсивной терапии, Медицинский и образовательный центр Эмама Хоссейна, Университет медицинских наук им. Шахида Бехешти, Тегеран, Иран.
b Кафедра клинической фармации, Факультет фармации, Университет медицинских наук им. Шахида Бехешти, Тегеран, Иран.
Шахида Бехешти, Тегеран, Иран.
c Отделение фармацевтической помощи, Медицинский и образовательный центр Эмама Хоссейна, Университет медицинских наук им. Шахида Бехешти, Тегеран, Иран.
* Автор, ответственный за переписку: Электронная почта: [email protected]; [email protected]
Поступила в январе 2018 г.; Принято в декабре 2018 г.0/), который разрешает неограниченное использование, распространение и воспроизведение на любом носителе при условии правильного цитирования оригинальной работы. Эта статья цитировалась в других статьях PMC.
Abstract
Взаимосвязь витамина D3 с продолжительностью искусственной вентиляции легких и смертностью до сих пор неизвестна. Таким образом, это исследование было направлено на определение влияния использования высоких доз витамина D на продолжительность ИВЛ среди пациентов, поступивших в отделение интенсивной терапии. Текущее двойное слепое клиническое исследование было проведено на 44 взрослых пациентах, находящихся на искусственной вентиляции легких. С помощью рандомизации переставленных блоков пациенты были набраны в группы вмешательства и плацебо. В группе плацебо четыре пациента были исключены из-за смерти до 72 часов. Уровень витамина D измеряли в обеих группах при поступлении и на 79011-й -й день исследования. В группах вмешательства и плацебо внутримышечно вводили 300 000 МЕ витамина D и такое же количество плацебо соответственно. Оценка по шкале SOFA и CPIS проводилась ежедневно в течение 7 дней, а также на 14 и 28 дней исследования.Также регистрировали продолжительность ИВЛ и летальность. В экспериментальную группу были включены 14 мужчин и 8 женщин, а в контрольную группу – 13 мужчин и 5 женщин. Достоверных различий в исходных характеристиках пациентов, включая пол и возраст, не было. Средняя продолжительность ИВЛ составила 17,63 ± 14 дней в группе вмешательства по сравнению с 27,72 ± 22,48 дня в контрольной группе ( p = 0,06). Смертность в контрольной и интервенционной группах составила 61.
С помощью рандомизации переставленных блоков пациенты были набраны в группы вмешательства и плацебо. В группе плацебо четыре пациента были исключены из-за смерти до 72 часов. Уровень витамина D измеряли в обеих группах при поступлении и на 79011-й -й день исследования. В группах вмешательства и плацебо внутримышечно вводили 300 000 МЕ витамина D и такое же количество плацебо соответственно. Оценка по шкале SOFA и CPIS проводилась ежедневно в течение 7 дней, а также на 14 и 28 дней исследования.Также регистрировали продолжительность ИВЛ и летальность. В экспериментальную группу были включены 14 мужчин и 8 женщин, а в контрольную группу – 13 мужчин и 5 женщин. Достоверных различий в исходных характеристиках пациентов, включая пол и возраст, не было. Средняя продолжительность ИВЛ составила 17,63 ± 14 дней в группе вмешательства по сравнению с 27,72 ± 22,48 дня в контрольной группе ( p = 0,06). Смертность в контрольной и интервенционной группах составила 61. 1% против 36,3% ( p = 0,00) соответственно. Введение высоких доз витамина D может снизить смертность у пациентов, находящихся на искусственной вентиляции легких. Несмотря на уменьшение продолжительности ИВЛ на 10 дней, разница не была статистически значимой. Рекомендуются более масштабные исследования.
1% против 36,3% ( p = 0,00) соответственно. Введение высоких доз витамина D может снизить смертность у пациентов, находящихся на искусственной вентиляции легких. Несмотря на уменьшение продолжительности ИВЛ на 10 дней, разница не была статистически значимой. Рекомендуются более масштабные исследования.
Ключевые слова: Витамин D, Успешное отлучение от груди, Искусственная вентиляция легких, Интенсивная терапия
Введение
Витамин D является жирорастворимым витамином, который вырабатывается в коже под действием УФ-излучения или поступает с пищей.Связь между дефицитом витамина D и повышенным риском смертности была отмечена у некритических пациентов (1–3). С момента первого сообщения о дефиците витамина D среди пациентов в критическом состоянии в 2009 году в нескольких исследованиях была показана высокая распространенность его дефицита у пациентов в отделениях интенсивной терапии, от 50 до 100 процентов (4-7).
У пациентов в критическом состоянии клинический исход дефицита витамина D и роль его добавок не ясны. В одном исследовании сообщалось, что дефицит витамина D напрямую связан с тяжестью заболевания и уровнем смертности, но может не приводить к увеличению продолжительности пребывания в отделении интенсивной терапии, увеличению частоты почечной недостаточности и сепсиса (8-10).В отличие от этих исследований, многие исследования показали, что дефицит витамина D имеет тесную связь с длительной госпитализацией в отделениях интенсивной терапии, повторной госпитализацией, сепсисом и смертностью (11-13). Некоторые исследования также показали, что дефицит витамина D также может увеличить риск дыхательной недостаточности (10, 14).
В одном исследовании сообщалось, что дефицит витамина D напрямую связан с тяжестью заболевания и уровнем смертности, но может не приводить к увеличению продолжительности пребывания в отделении интенсивной терапии, увеличению частоты почечной недостаточности и сепсиса (8-10).В отличие от этих исследований, многие исследования показали, что дефицит витамина D имеет тесную связь с длительной госпитализацией в отделениях интенсивной терапии, повторной госпитализацией, сепсисом и смертностью (11-13). Некоторые исследования также показали, что дефицит витамина D также может увеличить риск дыхательной недостаточности (10, 14).
Несмотря на некоторые исследования роли витамина D у пациентов в критическом состоянии, контролируемых исследований, оценивающих роль витамина D в продолжительности искусственной вентиляции легких, не проводилось.Таким образом, основной целью настоящего исследования было определить влияние добавок витамина D на продолжительность ИВЛ у пациентов ОИТ.
Experimental
Настоящее одноцентровое двойное слепое клиническое исследование было проведено на пациентах, поступивших в отделение интенсивной терапии больницы имама Хусейна после одобрения заместителем по исследованиям Университета медицинских наук Шахида Бехешти, Тегеран, Иран. Размер выборки оценивался в 44 человека в соответствии с предыдущими исследованиями и уравнением для расчета размера выборки (15).Пациенты были отобраны с использованием удобного метода выборки в соответствии с критериями включения, а затем разделены на две группы, вмешательство и плацебо, в соотношении 1: 1 путем рандомизации с перестановкой блоков. Участники этого исследования были набраны из взрослых (возраст от 18 до 65 лет) пациентов с механической вентиляцией легких. Критерии исключения: отказ законного опекуна пациента от участия в исследовании, смерть пациента менее чем через 72 ч после включения в исследование, нарушение функции почек (СКФ < 30 мл/мин), начало диализа во время лечения. исследование, прием витамина D в течение последних 15 дней, гипо/гиперкальциемия (скорректированный уровень кальция менее 8 мг/дл или выше 10), печеночная недостаточность (стадия C по Чайлд-Пафу), дисфункция паращитовидной железы, CPIS (клиническая шкала легочной инфекции). ) > 6, МНО > 1.5, тромбоциты < 80 000 и гемодинамические нарушения (САД менее 60 мм рт.
Размер выборки оценивался в 44 человека в соответствии с предыдущими исследованиями и уравнением для расчета размера выборки (15).Пациенты были отобраны с использованием удобного метода выборки в соответствии с критериями включения, а затем разделены на две группы, вмешательство и плацебо, в соотношении 1: 1 путем рандомизации с перестановкой блоков. Участники этого исследования были набраны из взрослых (возраст от 18 до 65 лет) пациентов с механической вентиляцией легких. Критерии исключения: отказ законного опекуна пациента от участия в исследовании, смерть пациента менее чем через 72 ч после включения в исследование, нарушение функции почек (СКФ < 30 мл/мин), начало диализа во время лечения. исследование, прием витамина D в течение последних 15 дней, гипо/гиперкальциемия (скорректированный уровень кальция менее 8 мг/дл или выше 10), печеночная недостаточность (стадия C по Чайлд-Пафу), дисфункция паращитовидной железы, CPIS (клиническая шкала легочной инфекции). ) > 6, МНО > 1.5, тромбоциты < 80 000 и гемодинамические нарушения (САД менее 60 мм рт. ст. в течение трех часов подряд). В первый день ИВЛ было получено 5 мл венозной крови для определения уровня 25-ОН витамина D. Затем пациенты в группах вмешательства и плацебо получали внутримышечно 300 000 МЕ витамина D и такое же количество плацебо соответственно. И лекарство, и идентичное плацебо были приготовлены фармацевтической компанией «Дарупахш», Тегеран, Иран. Медсестры и врачи не знали об исследовательских группах.
ст. в течение трех часов подряд). В первый день ИВЛ было получено 5 мл венозной крови для определения уровня 25-ОН витамина D. Затем пациенты в группах вмешательства и плацебо получали внутримышечно 300 000 МЕ витамина D и такое же количество плацебо соответственно. И лекарство, и идентичное плацебо были приготовлены фармацевтической компанией «Дарупахш», Тегеран, Иран. Медсестры и врачи не знали об исследовательских группах.
Для всех включенных в исследование пациентов оценка острого физиологического и хронического состояния здоровья II (APACHE II) определялась при включении в исследование. Шкала последовательной оценки органной недостаточности (SOFA) и шкала клинической легочной инфекции (CPIS) оценивались ежедневно до 7 дней, а затем на 14 и 28 день исследования. Уровень витамина D измеряли на 79011-й -й день исследования. Кроме того, в заранее разработанной форме для каждого пациента регистрировали уровень сознания (по шкале комы Гласкоу), продолжительность ИВЛ, длительность пребывания в отделении интенсивной терапии и 28-дневную смертность. Данные были проанализированы с помощью программного обеспечения SPSS версии 24 с использованием хи-квадрата, независимого t-критерия и повторных измерений ANOVA и критерия Уилкоксона. Колмогорова-Смирнова использовали для проверки нормальности распределения данных. Критерий Манна-Уитни применялся для непараметрических данных. В противном случае использовали независимый t-критерий.
Данные были проанализированы с помощью программного обеспечения SPSS версии 24 с использованием хи-квадрата, независимого t-критерия и повторных измерений ANOVA и критерия Уилкоксона. Колмогорова-Смирнова использовали для проверки нормальности распределения данных. Критерий Манна-Уитни применялся для непараметрических данных. В противном случае использовали независимый t-критерий.
Результаты
Всего в исследование было включено 44 пациента, из которых 4 пациента были исключены из исследования в связи со смертью менее чем через 72 ч, которые находились в контрольной группе. Остальные 40 пациентов включали 27 мужчин и 13 женщин (14 мужчин и 8 женщин в группе вмешательства).По критерию хи-квадрат между двумя группами не было существенной разницы по полу ( p = 0,55). Средний возраст составил 52 ± 22,1 и 56 ± 22,1 года в экспериментальной и контрольной группах соответственно ( p = 0,55). Кроме того, не было статистически значимых различий между исходными характеристиками пациентов в двух группах исследования (2).
Таблица 1
Исходные характеристики пациентов в контрольной и интервенционной группах исследования
| Переменные | Группы | р -значение | ||
|---|---|---|---|---|
| Контроль (n = 18) | Вмешательство (n = 22) | |||
| Возраст (год) | 56 ± 22.1 | 52 ± 22.1 | 0.55 | 9015 | 9 | 13 | 14 | 0. 55 55 |
| Оценка дивана Базовый | 6.6 ± 1,29 | 6.04 ± 0,95 | 0.85 | |
| CPI Оценка Baseline | 3.4 ± 1,6 | 3.13 ± 0, | 3,13 ± 0, | 19.77 ± 4.57 | 18.81 ± 4 | 0.49 |
| Vit D (NG / DL) Persone с дефицитом витамина D (<30 нг/дл) | 11.35 ± 18,23 | 8,43 ± 6,8 | 0,98 | |
Измерение уровня витамина D на седьмой день исследования показало увеличение с 8,43 ± 6,80 до 10,48 ± 9,80 нг/дл в группе вмешательства от 18,23 до 11,16 ± 18,22 нг/дл в контрольной группе, но разница между двумя группами не достигала значимого уровня ( p = 0,29).
Продолжительность ИВЛ и срок пребывания в стационаре составили 27,72 ± 22.48 против 17,63 ± 14,00 ( p = 0,06) и 19,50 ± 12,20 против 28,72 ± 23,58 ( p = 0,06) в контрольной и интервенционной группах исследования соответственно. Кроме того, уровень смертности в контрольной группе и группе вмешательства составил 61,1% против 36,3% ( p = 0,00) соответственно. Данные показаны в .
Таблица 2
Сравнение средней продолжительности госпитализации, продолжительности ИВЛ и смертности больных в двух группах
| Группа | р -значение | |||
|---|---|---|---|---|
| Контроль (n = 18) | Вмешательство (n = 22) | |||
| ИВЛ (дни) | 27. 72 ± 22.48 72 ± 22.48 | 17.63 ± 14 | 0.06 | 0,06 | Длина проживания ICU (дни) | 28.72 ± 23.58 | 19,5 ± 12.2 | 0,06 | 0,06 |
| Смертность (%) | 11 (61.1) | 8 (36,3) | 0,00 | |
Повторные измерения ANOVA использовали для оценки временного эффекта, который не показал достоверной разницы между и внутригрупповыми результатами по шкале SOFA ни в одной из ситуаций ( p = 0.13, р = 0,14). Кроме того, анализ ежедневных изменений показателя SOFA по сравнению с исходным уровнем с использованием повторных измерений ANOVA не показал существенных изменений в контрольной группе. Однако это сравнение выявило значительное снижение на 5 , 6 и 7 день исследования в группе вмешательства ( p -значения, равные 0,012, 0,006 и 0,002 соответственно). Данные показаны в .
Кроме того, анализ ежедневных изменений показателя SOFA по сравнению с исходным уровнем с использованием повторных измерений ANOVA не показал существенных изменений в контрольной группе. Однако это сравнение выявило значительное снижение на 5 , 6 и 7 день исследования в группе вмешательства ( p -значения, равные 0,012, 0,006 и 0,002 соответственно). Данные показаны в .
Изменения средних баллов по шкале SOFA в двух группах с течением времени
Изменения показателей CPIS с течением времени показаны на .Анализ ежедневных изменений CPIS по сравнению с исходным уровнем с использованием повторных измерений ANOVA показал, что средний балл значительно увеличился в контрольной группе (значения p составили 0,47, 0,10, 0,00, 0,01, 0,00, 0,00 и 0,00 соответственно), однако , те же анализы в группе вмешательства не достигли значимого уровня. Несмотря на разный характер изменений в двух группах исследования, частота культуральных бактериальных инфекций дыхательных путей не показала статистически значимой разницы ( p = 0. 74).
74).
Изменение средних показателей CPIS в двух группах с течением времени
Обсуждение
Результаты исследования показали, что среднее количество дней ИВЛ уменьшилось с 27 до 17 дней в группе витамина D, но это не достигло статистической значимости . Возможно, дефицит витамина D может быть одним из виновников зависимости от аппарата ИВЛ и может продлить время отлучения от аппарата ИВЛ. В исследовании Quraishi et al. , измерили уровни 25-гидроксивитамина D в плазме у тяжелобольных хирургических пациентов при поступлении в ОИТ и пришли к выводу, что уровни 25-гидроксивитамина D обратно пропорциональны продолжительности респираторной поддержки (16).
Другим интересным открытием текущего исследования было влияние высоких доз витамина D на смертность пациентов от всех причин. Наши результаты показали, что количество выживших пациентов было значительно выше в группе вмешательства, чем в контроле. Таким образом, инъекции высоких доз витамина D могут увеличить выживаемость пациентов.
Адекватные уровни витамина D необходимы для регуляции функции иммунной системы, а его дефицит приводит к нарушению иммунной функции.Это может привести к повышенному риску инфекций, особенно пневмонии, связанной с ИВЛ, системного воспаления и синдромов полиорганной дисфункции, что может увеличить уровень смертности и продолжительность пребывания в отделении интенсивной терапии (17, 18).
Результаты приема витамина D противоречивы. В исследовании Aygencel et al. сообщили, что уровень смертности был значительно выше в группе с недостатком витамина D по сравнению с группой с достаточным количеством витамина D, но дефицит витамина D в этом исследовании не был независимым фактором риска смертности (19).В другом исследовании Putzu et al. показало, что прием витамина D у пациентов в критическом состоянии может снизить смертность без значительных побочных эффектов. Кроме того, Miroliae, et al., показали, что добавление витамина D у пациентов с вентилятор-ассоциированной пневмонией может значительно снизить уровень прокальцитонина (20). Кроме того, наши результаты показали, что добавление витамина D может снизить уровень смертности и продолжительность пребывания в отделении интенсивной терапии. В отличие от наших результатов Langlois et al., в систематическом обзоре сделан вывод о том, что введение витамина D не улучшает клинические исходы (7).
Кроме того, наши результаты показали, что добавление витамина D может снизить уровень смертности и продолжительность пребывания в отделении интенсивной терапии. В отличие от наших результатов Langlois et al., в систематическом обзоре сделан вывод о том, что введение витамина D не улучшает клинические исходы (7).
В нашем исследовании, несмотря на одну инъекцию 300 000 единиц витамина D внутримышечно, уровень 25-OH витамина D значительно не увеличился через 7 дней после введения. Амрейн и др. показали, что прием высоких пероральных доз у пациентов с дефицитом витамина D может улучшить его уровень в крови в течение 2 дней. В этом исследовании пациентам в группе вмешательства было назначено 540 000 единиц пероральных препаратов, что превышало дозу нашего исследования.В отличие от наших результатов, они обнаружили повышение уровня витамина D выше 25 нг/дл, что не соответствует результатам этого исследования. Однако влияние его улучшения на прогноз пациентов не оценивалось (21). В другом исследовании Gorman et al. показал, что у субъектов, получавших однократную внутримышечную инъекцию высокой дозы витамина D, уровни 25-OH витамина D в сыворотке повышались через 3, 4 и 24 недели после инъекции, достигая пика через 4 недели. Этот вывод согласуется с нашими результатами, которые не показали повышения уровня витамина D в сыворотке через неделю после инъекции (22).
В другом исследовании Gorman et al. показал, что у субъектов, получавших однократную внутримышечную инъекцию высокой дозы витамина D, уровни 25-OH витамина D в сыворотке повышались через 3, 4 и 24 недели после инъекции, достигая пика через 4 недели. Этот вывод согласуется с нашими результатами, которые не показали повышения уровня витамина D в сыворотке через неделю после инъекции (22).
Заключение
Это исследование показало, что введение высоких доз витамина D может эффективно снижать продолжительность искусственной вентиляции легких, продолжительность госпитализации и уровень смертности у пациентов в ОИТ. Для подтверждения этих выводов необходимы дальнейшие исследования с большим размером выборки и многоцентровым дизайном.
Благодарность
Исследователи хотели бы выразить благодарность заместителю по исследованиям Университета медицинских наук им. Шахида Бехешти, Тегеран, Иран, и персоналу отделения интенсивной терапии больницы имама Хусейна.
Ссылки
1. Белакович Г., Глууд Л.Л., Николова Д., Уитфилд К., Веттерслев Дж., Симонетти Р.Г., Белакович М., Глууд С. Добавка витамина D для предотвращения смертности у взрослых. Кокрановская система баз данных. Версия 2014: CD007470. [PubMed] [Google Scholar]2. Dobnig H, Pilz S, Scharnagl H, Renner W, Seelhorst U, Wellnitz B, Kinkeldei J, Boehm BO, Weihrauch G, Maerz W. Независимая связь низких уровней 25-гидроксивитамина d и 1,25-дигидроксивитамина d в сыворотке со всеми причин и смертности от сердечно-сосудистых заболеваний.Арка Стажер Мед. 2008; 168:1340–9. [PubMed] [Google Scholar]3. Zittermann A, Iodice S, Pilz S, Grant WB, Bagnardi V, Gandini S. Дефицит витамина D и риск смертности среди населения в целом: метаанализ проспективных когортных исследований. Являюсь. Дж. Клин. Нутр. 2012;95:91–100. [PubMed] [Google Scholar]4. Ли П., Эйсман Дж. А., Центр Дж. Р. Дефицит витамина D у пациентов в критическом состоянии. Н. англ. Дж. Мед. 2009; 360:1912–4. [PubMed] [Google Scholar]5. Азим А., Ахмед А., Ядав С., Барония А.К., Гурджар М., Годбол М.М., Поддар Б., Сингх Р.К.Распространенность дефицита витамина D у пациентов в критическом состоянии и его влияние на исход: опыт центра третичной медицинской помощи в Северной Индии (обсервационное исследование) J. Intensive Care. 2013;1 [Бесплатная статья PMC] [PubMed] [Google Scholar]6. Sauneuf B, Brunet J, Lucidarme O, du Cheyron D. Распространенность и факторы риска дефицита витамина D у пациентов в критическом состоянии. Воспаление. Цели лекарств от аллергии. 2013;12:223–9. [PubMed] [Google Scholar]7. Ланглуа П.Л., Швец К., Д’Арагон Ф., Хейланд Д.К., Мансанарес В.Добавление витамина D в критических состояниях: систематический обзор и метаанализ. клин. Нутр. 2018;37:1238–46. [PubMed] [Google Scholar]8. Перрон Р.М., Ли П. Эффективность приема высоких доз витамина D у пациентов в критическом состоянии. Воспаление. Цели лекарств от аллергии. 2013;12:273–81. [PubMed] [Google Scholar]9. Браун А.Б., Литонхуа А.А., Моромизато Т.
Азим А., Ахмед А., Ядав С., Барония А.К., Гурджар М., Годбол М.М., Поддар Б., Сингх Р.К.Распространенность дефицита витамина D у пациентов в критическом состоянии и его влияние на исход: опыт центра третичной медицинской помощи в Северной Индии (обсервационное исследование) J. Intensive Care. 2013;1 [Бесплатная статья PMC] [PubMed] [Google Scholar]6. Sauneuf B, Brunet J, Lucidarme O, du Cheyron D. Распространенность и факторы риска дефицита витамина D у пациентов в критическом состоянии. Воспаление. Цели лекарств от аллергии. 2013;12:223–9. [PubMed] [Google Scholar]7. Ланглуа П.Л., Швец К., Д’Арагон Ф., Хейланд Д.К., Мансанарес В.Добавление витамина D в критических состояниях: систематический обзор и метаанализ. клин. Нутр. 2018;37:1238–46. [PubMed] [Google Scholar]8. Перрон Р.М., Ли П. Эффективность приема высоких доз витамина D у пациентов в критическом состоянии. Воспаление. Цели лекарств от аллергии. 2013;12:273–81. [PubMed] [Google Scholar]9. Браун А.Б., Литонхуа А.А., Моромизато Т. , Гиббонс Ф.К., Джованнуччи Э., Кристофер К.Б. Связь низкого уровня 25-гидроксивитамина D в сыворотке крови с острым повреждением почек у больных в критическом состоянии. крит.Уход Мед. 2012;40:3170–9. [PubMed] [Google Scholar] 10. Моромизато Т., Литонхуа А.А., Браун А.Б., Гиббонс Ф.К., Джованнуччи Э., Кристофер К.Б. Связь низкого уровня 25-гидроксивитамина D в сыворотке крови и сепсиса у больных в критическом состоянии. крит. Уход Мед. 2014;42:97–107. [PubMed] [Google Scholar] 11. McNally JD, Menon K, Chakraborty P, Fisher L, Williams KA, Al-Dirbashi OY, Doherty DR. Связь статуса витамина D с педиатрическими критическими заболеваниями. Педиатрия. 2012; 130:429–36. [PubMed] [Google Scholar] 12. Кемпкер Дж. А., Хан Дж. Э., Тангприча В., Зиглер Т. Р., Мартин Г. С.Витамин D и сепсис: новые отношения. Дерматоэндокринол. 2012;4:101–8. [Бесплатная статья PMC] [PubMed] [Google Scholar]13. Кемпкер Дж. А., Вест К. Г., Кемпкер Р. Р., Сивамогсатхам О., Альварес Дж. А., Тангприча В., Зиглер Т. Р., Мартин Г.
, Гиббонс Ф.К., Джованнуччи Э., Кристофер К.Б. Связь низкого уровня 25-гидроксивитамина D в сыворотке крови с острым повреждением почек у больных в критическом состоянии. крит.Уход Мед. 2012;40:3170–9. [PubMed] [Google Scholar] 10. Моромизато Т., Литонхуа А.А., Браун А.Б., Гиббонс Ф.К., Джованнуччи Э., Кристофер К.Б. Связь низкого уровня 25-гидроксивитамина D в сыворотке крови и сепсиса у больных в критическом состоянии. крит. Уход Мед. 2014;42:97–107. [PubMed] [Google Scholar] 11. McNally JD, Menon K, Chakraborty P, Fisher L, Williams KA, Al-Dirbashi OY, Doherty DR. Связь статуса витамина D с педиатрическими критическими заболеваниями. Педиатрия. 2012; 130:429–36. [PubMed] [Google Scholar] 12. Кемпкер Дж. А., Хан Дж. Э., Тангприча В., Зиглер Т. Р., Мартин Г. С.Витамин D и сепсис: новые отношения. Дерматоэндокринол. 2012;4:101–8. [Бесплатная статья PMC] [PubMed] [Google Scholar]13. Кемпкер Дж. А., Вест К. Г., Кемпкер Р. Р., Сивамогсатхам О., Альварес Дж. А., Тангприча В., Зиглер Т. Р., Мартин Г. С. Статус витамина D и риск внутрибольничных инфекций у взрослых в критическом состоянии: проспективное когортное исследование. Плос один. 2015;10:e0122136. [Бесплатная статья PMC] [PubMed] [Google Scholar]14. Тикетт Д.Р., Моромизато Т., Литонджуа А.А., Амрейн К., Кураиши С.А., Ли-Сарвар К.А., Могенсен К.М., Пёртл С.В., Гиббонс Ф.К., Камарго К.А. младший, Джованнуччи Э., Кристофер К.Б.Связь между догоспитальным статусом витамина D и возникновением острой дыхательной недостаточности у пациентов в критическом состоянии: ретроспективное когортное исследование. Дыхание BMJ открытое. Рез. 2015;2:e000074. [Бесплатная статья PMC] [PubMed] [Google Scholar]15. Хан Дж.Э., Джонс Дж.Л., Тангприча В., Браун М.А., Браун Л.А.С., Хао Л., Хеббар Г., Ли М.Дж., Лю С., Зиглер Т.Р., Мартин Г.С. Введение высоких доз витамина D пациентам отделения интенсивной терапии с вентиляцией легких: пилотное двойное слепое рандомизированное контролируемое исследование. Дж. Клин. Перевод Эндокринол. 2016; 4:59–65.[Бесплатная статья PMC] [PubMed] [Google Scholar]16.
С. Статус витамина D и риск внутрибольничных инфекций у взрослых в критическом состоянии: проспективное когортное исследование. Плос один. 2015;10:e0122136. [Бесплатная статья PMC] [PubMed] [Google Scholar]14. Тикетт Д.Р., Моромизато Т., Литонджуа А.А., Амрейн К., Кураиши С.А., Ли-Сарвар К.А., Могенсен К.М., Пёртл С.В., Гиббонс Ф.К., Камарго К.А. младший, Джованнуччи Э., Кристофер К.Б.Связь между догоспитальным статусом витамина D и возникновением острой дыхательной недостаточности у пациентов в критическом состоянии: ретроспективное когортное исследование. Дыхание BMJ открытое. Рез. 2015;2:e000074. [Бесплатная статья PMC] [PubMed] [Google Scholar]15. Хан Дж.Э., Джонс Дж.Л., Тангприча В., Браун М.А., Браун Л.А.С., Хао Л., Хеббар Г., Ли М.Дж., Лю С., Зиглер Т.Р., Мартин Г.С. Введение высоких доз витамина D пациентам отделения интенсивной терапии с вентиляцией легких: пилотное двойное слепое рандомизированное контролируемое исследование. Дж. Клин. Перевод Эндокринол. 2016; 4:59–65.[Бесплатная статья PMC] [PubMed] [Google Scholar]16.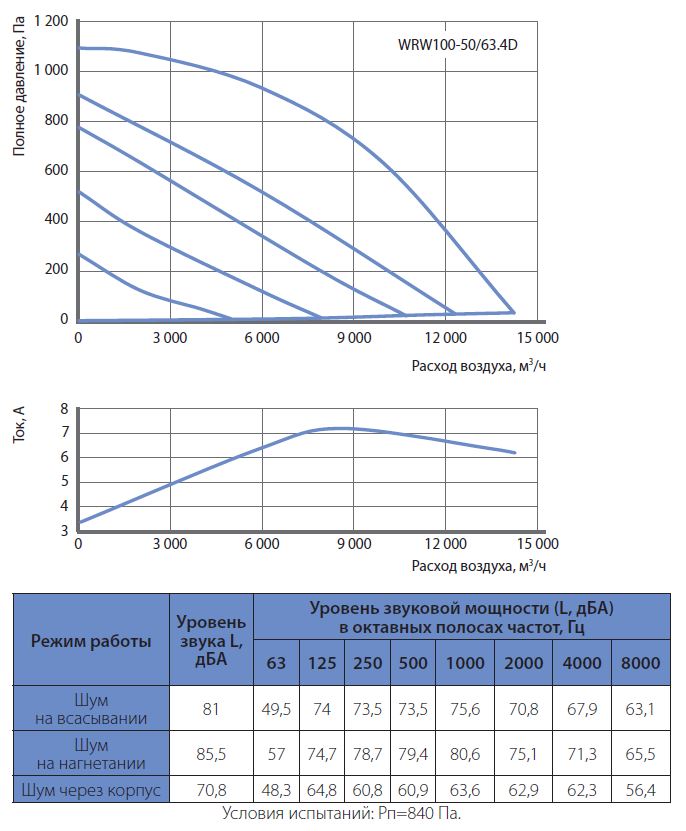 Quraishi SA, McCarthy C, Blum L, Cobb JP, Camargo CA Jr. Уровни 25-гидроксивитамина D в плазме при начале лечения и продолжительности искусственной вентиляции легких у тяжелобольных хирургических пациентов. JPEN Дж. Парентер. Энтераль. Нутр. 2016;40:273–8. [Бесплатная статья PMC] [PubMed] [Google Scholar]17. Дженг Л., Ямщиков А.В., Джадд С.Е., Блумберг Х.М., Мартин Г.С., Циглер Т.Р., Тангприча В. Изменения статуса витамина D и уровней антимикробных пептидов у пациентов в отделении интенсивной терапии с сепсисом.Дж. Пер. Мед. 2009;7:28. [Бесплатная статья PMC] [PubMed] [Google Scholar]18. Baeke F, Takiishi T, Korf H, Gysemans C, Mathieu C. Витамин D: модулятор иммунной системы. Курс. мнение Фармакол. 2010;10:482–96. [PubMed] [Google Scholar] 19. Aygencel G, Turkoglu M, Tuncel AF, Candir BA, Bildaci YD, Pasaoglu H. Связана ли недостаточность витамина D со смертностью пациентов в критическом состоянии? крит. Уход Рез. Практика. 2013;2013:856747. [Бесплатная статья PMC] [PubMed] [Google Scholar]20.
Quraishi SA, McCarthy C, Blum L, Cobb JP, Camargo CA Jr. Уровни 25-гидроксивитамина D в плазме при начале лечения и продолжительности искусственной вентиляции легких у тяжелобольных хирургических пациентов. JPEN Дж. Парентер. Энтераль. Нутр. 2016;40:273–8. [Бесплатная статья PMC] [PubMed] [Google Scholar]17. Дженг Л., Ямщиков А.В., Джадд С.Е., Блумберг Х.М., Мартин Г.С., Циглер Т.Р., Тангприча В. Изменения статуса витамина D и уровней антимикробных пептидов у пациентов в отделении интенсивной терапии с сепсисом.Дж. Пер. Мед. 2009;7:28. [Бесплатная статья PMC] [PubMed] [Google Scholar]18. Baeke F, Takiishi T, Korf H, Gysemans C, Mathieu C. Витамин D: модулятор иммунной системы. Курс. мнение Фармакол. 2010;10:482–96. [PubMed] [Google Scholar] 19. Aygencel G, Turkoglu M, Tuncel AF, Candir BA, Bildaci YD, Pasaoglu H. Связана ли недостаточность витамина D со смертностью пациентов в критическом состоянии? крит. Уход Рез. Практика. 2013;2013:856747. [Бесплатная статья PMC] [PubMed] [Google Scholar]20. Миролиаи А.Э., Саламзаде Дж., Шокоуи С., Фатеми А., Ардехали С.Х., Хаджисмаили М., Сахраи З.Влияние добавок витамина D на прокальцитонин как прогностический биомаркер у пациентов с вентилятор-ассоциированной пневмонией, осложненной дефицитом витамина D. Иран. Дж. Фарм. Рез. 2017;16:1254–63. [Бесплатная статья PMC] [PubMed] [Google Scholar]21. Amrein K, Sourij H, Wagner G, Holl A, Pieber TR, Smolle KH, Stojakovic T, Schnedl C, Dobnig H. Кратковременные эффекты перорального приема высоких доз витамина D3 у пациентов с дефицитом витамина D в критическом состоянии: рандомизированный, двойной -слепое плацебо-контролируемое пилотное исследование.крит. Уход. 2011;15:R104. [Бесплатная статья PMC] [PubMed] [Google Scholar]22. Горман С., Зафирау М.З., Лим Э.М., Кларк М.В., Дхамрайт Г., Флери Н., Уолш Д.П., Кауфманн М., Джонс Г., Лукас Р.М. Высокие дозы витамина D внутримышечно обеспечивают длительное умеренное повышение уровня 25-гидроксивитамина D в сыворотке и кратковременные изменения уровня кальция в плазме.
Миролиаи А.Э., Саламзаде Дж., Шокоуи С., Фатеми А., Ардехали С.Х., Хаджисмаили М., Сахраи З.Влияние добавок витамина D на прокальцитонин как прогностический биомаркер у пациентов с вентилятор-ассоциированной пневмонией, осложненной дефицитом витамина D. Иран. Дж. Фарм. Рез. 2017;16:1254–63. [Бесплатная статья PMC] [PubMed] [Google Scholar]21. Amrein K, Sourij H, Wagner G, Holl A, Pieber TR, Smolle KH, Stojakovic T, Schnedl C, Dobnig H. Кратковременные эффекты перорального приема высоких доз витамина D3 у пациентов с дефицитом витамина D в критическом состоянии: рандомизированный, двойной -слепое плацебо-контролируемое пилотное исследование.крит. Уход. 2011;15:R104. [Бесплатная статья PMC] [PubMed] [Google Scholar]22. Горман С., Зафирау М.З., Лим Э.М., Кларк М.В., Дхамрайт Г., Флери Н., Уолш Д.П., Кауфманн М., Джонс Г., Лукас Р.М. Высокие дозы витамина D внутримышечно обеспечивают длительное умеренное повышение уровня 25-гидроксивитамина D в сыворотке и кратковременные изменения уровня кальция в плазме. J. АОАС междунар. 2017; 100:1337–44. [PubMed] [Google Scholar]
J. АОАС междунар. 2017; 100:1337–44. [PubMed] [Google Scholar]%PDF-1.3 % 39 0 объект >/OCGs[83 0 R]>>/OpenAction[40 0 R/Fit]/PageLabels 34 0 R/PageMode/UseNone/Pages 36 0 R/Type/Catalog>> эндообъект 150 0 объект >/Шрифт>>>/Поля 151 0 R>> эндообъект 37 0 объект >поток 2015-02-20T12:43:48Z2015-02-23T15:44:57+05:30XPP2015-02-23T15:44:57+05:30приложение/pdf
UUID: fe10ecfc-8b88-49cc-b7d8-452b38f23c6buuid: f883b4f9-a770-4020-a593-084af4efbb37 конечный поток эндообъект 34 0 объект > эндообъект 36 0 объект > эндообъект 40 0 объект >/Font>/ProcSet[/PDF/Text]/XObject>>>/Rotate 0/TrimBox[9 9 594 792]/Type/Page>> эндообъект 1 0 объект >/Font>/ProcSet[/PDF/Text]/XObject>>>/Rotate 0/TrimBox[9 9 594 792]/Type/Page>> эндообъект 4 0 объект >/Font>/ProcSet[/PDF/Text]/XObject>>>/Rotate 0/TrimBox[9 9 594 792]/Type/Page>> эндообъект 7 0 объект >/ExtGState>/Font>/ProcSet[/PDF/Text/ImageC/ImageI]/XObject>>>/Rotate 0/TrimBox[9 9 594 792]/Type/Page>> эндообъект 16 0 объект >/Font>/ProcSet[/PDF/Text]/XObject>>>/Rotate 0/TrimBox[9 9 594 792]/Type/Page>> эндообъект 19 0 объект >/Font>/ProcSet[/PDF/Text]/XObject>>>/Rotate 0/TrimBox[9 9 594 792]/Type/Page>> эндообъект 22 0 объект >/Font>/ProcSet[/PDF/Text]/XObject>>>/Rotate 0/TrimBox[9 9 594 792]/Type/Page>> эндообъект 149 0 объект >поток HWmo6n!f”YRSpm7/KZ0i%~Geه%Ht/E_;W3Ŝe4U’j8@!)>x\>7[შjrfSRj̎nGN=ZN9VA%v(ԥZRXa2#xLl&WBWjg+[% *d!tzʟDjԙ]/`qνjeZ$e3K] W%A&kmE (Q&F4QsN=y^eU:署*ҥTv49@:/E]8(ҙ8 Έ’UHRZ0zn5ee͕L2]>ĞXCF2yR_C;?1ϼ_[aY*6w %Y&Ҏ4BҪ536SaßBUПроизошла ошибка при настройке пользовательского файла cookie
Этот сайт использует файлы cookie для повышения производительности. Если ваш браузер не принимает файлы cookie, вы не можете просматривать этот сайт.
Если ваш браузер не принимает файлы cookie, вы не можете просматривать этот сайт.
Настройка браузера на прием файлов cookie
Существует множество причин, по которым файл cookie не может быть установлен правильно. Ниже приведены наиболее распространенные причины:
- В вашем браузере отключены файлы cookie. Вам необходимо сбросить настройки браузера, чтобы принять файлы cookie, или спросить вас, хотите ли вы принимать файлы cookie.
- Ваш браузер спрашивает, хотите ли вы принимать файлы cookie, и вы отказались.Чтобы принять файлы cookie с этого сайта, нажмите кнопку «Назад» и примите файл cookie.
- Ваш браузер не поддерживает файлы cookie. Попробуйте другой браузер, если вы подозреваете это.
- Дата на вашем компьютере в прошлом. Если часы вашего компьютера показывают дату до 1 января 1970 г.,
браузер автоматически забудет файл cookie. Чтобы это исправить, установите правильное время и дату на своем компьютере.
- Вы установили приложение, которое отслеживает или блокирует установку файлов cookie.Вы должны отключить приложение при входе в систему или проконсультироваться с системным администратором.

Почему этому сайту требуются файлы cookie?
Этот сайт использует файлы cookie для повышения производительности, запоминая, что вы вошли в систему, когда переходите со страницы на страницу. Предоставить доступ без файлов cookie потребует от сайта создания нового сеанса для каждой посещаемой вами страницы, что замедляет работу системы до неприемлемого уровня.
Что сохраняется в файле cookie?
Этот сайт не хранит ничего, кроме автоматически сгенерированного идентификатора сеанса в файле cookie; никакая другая информация не фиксируется.
Как правило, в файле cookie может храниться только та информация, которую вы предоставляете, или выбор, который вы делаете при посещении веб-сайта. Например, сайт
не может определить ваше имя электронной почты, если вы не решите ввести его. Разрешение веб-сайту создавать файлы cookie не дает этому или любому другому сайту доступ к
остальной части вашего компьютера, и только сайт, создавший файл cookie, может его прочитать.
Например, сайт
не может определить ваше имя электронной почты, если вы не решите ввести его. Разрешение веб-сайту создавать файлы cookie не дает этому или любому другому сайту доступ к
остальной части вашего компьютера, и только сайт, создавший файл cookie, может его прочитать.
| 1 шт.btw | ||
| 2 шт. | € 319,00 / € 638,00 btw | |
| 3 шт. btw | ||
| 4 шт. | € 319,00 / 1 276,00 € кстати | |
| 5 штук | € 319,00 / € 1.595,00 € 385,99 / € 1.929,95 вкл. btw | |
| 6 штук | 319,00 € / 1 914,00 € btw | |
| 7 шт. | 319,00 € / 2 233,00 € btw | |
| 8 шт. | € 319,00 / € 2.552,00 кстати | |
| 9 штук | € 319,00 / € 2. € 385,99 / € 3.473,91 вкл. btw | |
| 10 штук | € 319,00 / € 3.190,00 btw | |
| 11 шт. | € 319,00 / € 3.509,00 btw | |
| 12 штук | € 319,00 / € 3.828,00 кстати | |
| 13 штук | € 319,00 / € 4.147,00 € 385,99 / € 5.017,87 вкл. btw | |
| 14 штук | € 319,00 / 4 466,00 € btw | |
| 15 штук | € 319,00 / € 4.785,00 btw | |
| 16 шт. | € 319,00 / € 5.104,00 кстати | |
| 17 штук | € 319,00 / € 5.423,00 € 385,99 / € 6.561,83 вкл. btw | |
| 18 штук | € 319,00 / € 5.742,00 btw | |
| 19 штук | € 319,00 / 6,061,00 € btw | |
20 шт. | € 319,00 / 6 380,00 € кстати | |
| 21 штукс | € 319,00 / € 6.699,00 € 385,99 / € 8.105,79 вкл. btw | |
| 22 шт. | € 319,00 / € 7.018,00 btw | |
| 23 шт. | € 319,00 / € 7.337,00 btw | |
| 24 шт. | € 319,00 / € 7.656,00 кстати | |
| 25 штук | € 319,00 / € 7.975,00 € 385,99 / € 9.649,75 вкл. btw | |
| 26 штук | € 319,00 / € 8.294,00 btw | |
| 27 штук | € 319,00 / 8 613,00 € btw | |
| 28 штук | € 319,00 / 8 932,00 € кстати | |
| 29 штук | € 319,00 / € 9.251,00 € 385,99 / € 11.193,71 вкл. btw | |
| 30 штук | € 319,00 / 9 570,00 € btw | |
31 шт. | 319,00 €/ 9 889,00 € 385,99 € / 69,95 € вкл. btw | |
| 32 шт. | € 319,00 / € 10.208,00 Кстати, | |
| 33 шт. | € 319,00 / € 10.527,00 € 385,99 / € 12.737,67 вкл. btw | |
| 34 шт. btw | ||
| 35 штук | 319,00 € / 11 165,00 € btw | |
| 36 шт. кстати | ||
| 37 штук | € 319,00 / € 11.803,00 € 385,99 / € 14.281,63 вкл. btw | |
| 38 штук | € 319,00 / € 12.122,00 € 385,99 / вкл. € 14,667 btw | |
| 39 шт. | € 319,00 / € 12.441,00 btw | |
| 40 штук | € 319,00 / € 12.760,00 кстати | |
| 41 штукс | € 319,00 / € 13.079,00 € 385,99 / € 15. | |
| 42 шт. | € 319,00 / € 13.398,00 btw | |
| 43 шт. | 319,00 € / 13.717,00 € btw | |
| 44 шт. | € 319,00 / € 14.036,00 кстати | |
| 45 штук | € 319,00 / € 14.355,00 € 385,99 / € 17.369,55 вкл. btw | |
| 46 шт. | € 319,00 / € 14.674,00 btw | |
| 47 штук | € 319,00 / 14 993,00 € € 385,99 / вкл. 18,141 € btw | |
| 48 штук | € 319,00 / 15 312,00 € кстати | |
| 49 штук | € 319,00 / € 15.631,00 € 385,99 / € 18.913,51 вкл. btw | |
| 50 штук | € 319,00 / € 15.950,00 кстати |
 871,00
871,00 825,59 вкл. btw
825,59 вкл. btwГраницы | Метаболический вклад альфапротеобактериального эндосимбионта в Apicomplexan Cardiosporidium cionae
Введение
Apicomplexa включает множество высоковирулентных патогенных организмов, таких как Plasmodium falciparum , Cryptosporidium parvum и Toxoplasma gondii , возбудителей малярии, криптоспоридиоза и токсоплазмоза соответственно. Малярия ежегодно уносит около полумиллиона человеческих жизней (Center for Disease Control, 2019), T. gondii , по оценкам, заражает до 60% населения большей части Европы (Pappas et al., 2009), а криптоспоридиоз ежегодно вызывает 3–5 миллионов случаев желудочно-кишечных заболеваний у детей только в Африке и Индии (Sow et al., 2016). Эти организмы представляют собой серьезную проблему для здоровья человека, но в результате наше понимание этого типа в значительной степени основано на небольшом подмножестве клинически значимых апикомплексов.Каждое многоклеточное животное, вероятно, является хозяином по крайней мере одного апикомплекса (Morrison, 2009), и это, вероятно, недооценка, поскольку многие виды могут содержать несколько видов апикомплекса. Apicomplexans были описаны у широкого круга позвоночных от птиц до морских млекопитающих (Jeurissen et al., 1996; Conrad et al., 2005), а также у книдарий (Kwong et al., 2019), моллюсков (Dyson et al. , 1993; Suja et al., 2016), членистоногих (Alarcón et al.
Малярия ежегодно уносит около полумиллиона человеческих жизней (Center for Disease Control, 2019), T. gondii , по оценкам, заражает до 60% населения большей части Европы (Pappas et al., 2009), а криптоспоридиоз ежегодно вызывает 3–5 миллионов случаев желудочно-кишечных заболеваний у детей только в Африке и Индии (Sow et al., 2016). Эти организмы представляют собой серьезную проблему для здоровья человека, но в результате наше понимание этого типа в значительной степени основано на небольшом подмножестве клинически значимых апикомплексов.Каждое многоклеточное животное, вероятно, является хозяином по крайней мере одного апикомплекса (Morrison, 2009), и это, вероятно, недооценка, поскольку многие виды могут содержать несколько видов апикомплекса. Apicomplexans были описаны у широкого круга позвоночных от птиц до морских млекопитающих (Jeurissen et al., 1996; Conrad et al., 2005), а также у книдарий (Kwong et al., 2019), моллюсков (Dyson et al. , 1993; Suja et al., 2016), членистоногих (Alarcón et al. , 2017; Criado-Fornelio et al., 2017) и хордовых (Ciancio et al., 2008; Саффо и др., 2010). Диапазон их хозяев огромен, а их разнообразие и приспособленность к паразитическому образу жизни не имеют себе равных.
, 2017; Criado-Fornelio et al., 2017) и хордовых (Ciancio et al., 2008; Саффо и др., 2010). Диапазон их хозяев огромен, а их разнообразие и приспособленность к паразитическому образу жизни не имеют себе равных.
Долгая история эволюции и адаптации к жизни внутри хозяина привела к ряду характерных геномных потерь и эволюции специализированных клеточных механизмов у апикомплексов (McFadden and Waller, 1997; Soldati et al., 2001; Roos, 2005; Janouskovec and Keeling, 2016; Frénal et al., 2017). Конкретные структурные адаптации этих организмов включают те, которые связаны с функциями, связанными с инфекцией и персистенцией хозяина; а именно остаток пластиды (апикопласт) и апикальный комплекс (McFadden and Waller, 1997; Soldati et al., 2001). Геномная редукция, связанная с паразитизмом у апикомплексов, включает потерю семейств генов для биосинтеза пуринов, аминокислот, стеролов, различных кофакторов, глиоксилатного цикла, компонентов эндомембраны и генов, связанных с подвижностью (Woo et al. , 2015; Janouskovec and Keeling, 2016). Кроме того, апикомплексаны также демонстрируют экспансию в семействах генов, связанных с инфекцией и персистенцией в клетках-хозяевах (Janouskovec and Keeling, 2016). Однако предположение о том, что эти геномные сигнатуры связаны с паразитизмом, основано на ограниченной информации, поскольку прямое сравнение с близкородственными свободноживущими сестринскими таксонами невозможно, а также отсутствуют известные свободноживущие апикомплексаны (Janouskovec and Keeling, 2016). .Однако доступны геномные данные фотосинтезирующих хромрид (Woo et al., 2015), которые, вероятно, отделились от апикомплексов 600–800 миллионов лет назад (Votýpka et al., 2016).
, 2015; Janouskovec and Keeling, 2016). Кроме того, апикомплексаны также демонстрируют экспансию в семействах генов, связанных с инфекцией и персистенцией в клетках-хозяевах (Janouskovec and Keeling, 2016). Однако предположение о том, что эти геномные сигнатуры связаны с паразитизмом, основано на ограниченной информации, поскольку прямое сравнение с близкородственными свободноживущими сестринскими таксонами невозможно, а также отсутствуют известные свободноживущие апикомплексаны (Janouskovec and Keeling, 2016). .Однако доступны геномные данные фотосинтезирующих хромрид (Woo et al., 2015), которые, вероятно, отделились от апикомплексов 600–800 миллионов лет назад (Votýpka et al., 2016).
Несмотря на высокую патогенность и паразитарную адаптацию многих членов, возникли вопросы о том, является ли Apicomplexa полностью паразитарной группой. Хотя это мнение уже давно упоминается в публикациях (Roos, 2005; Morrison, 2009; Gubbels and Duraisingh, 2012; Woo et al., 2015; Votýpka et al. , 2016; Макфадден и Йе, 2017 г .; Mathur et al., 2018), текущие данные свидетельствуют о том, что взаимодействия между апикомплексами и их хозяевами гораздо более разнообразны, чем предполагалось ранее. На самом деле вполне вероятно, что апикомплексы охватывают весь спектр от паразитизма до комменсализма и даже мутуализма (Saffo et al., 2010; Kwong et al., 2019; Rueckert et al., 2019). Однако то, что определяет границы этого континуума симбиотической ассоциации, до сих пор является предметом многочисленных споров (Ewald, 1987; Leung and Poulin, 2008; Johnson and Oelmüller, 2009).Филогенетический анализ показывает, что Nephromyces является сестрой клады гематозоев и тесно связан с высоковирулентными родами, такими как Plasmodium, Theiliera и Babesia (Muñoz-Gómez et al., 2019a). К настоящему времени в ранневетвящихся группах, таких как Gregarina и Corallicods, были обнаружены апикомплексные виды с изменчивыми жизненными стратегиями. Однако существование этого, как сообщается, мутуалистического таксона глубоко внутри Apicomplexa, сестры группы высоковирулентных паразитов крови, предполагает, что уникальная биология Nephromycidae может быть ответственна за такой переход к комменсальной или мутуалистической жизненной стратегии.
, 2016; Макфадден и Йе, 2017 г .; Mathur et al., 2018), текущие данные свидетельствуют о том, что взаимодействия между апикомплексами и их хозяевами гораздо более разнообразны, чем предполагалось ранее. На самом деле вполне вероятно, что апикомплексы охватывают весь спектр от паразитизма до комменсализма и даже мутуализма (Saffo et al., 2010; Kwong et al., 2019; Rueckert et al., 2019). Однако то, что определяет границы этого континуума симбиотической ассоциации, до сих пор является предметом многочисленных споров (Ewald, 1987; Leung and Poulin, 2008; Johnson and Oelmüller, 2009).Филогенетический анализ показывает, что Nephromyces является сестрой клады гематозоев и тесно связан с высоковирулентными родами, такими как Plasmodium, Theiliera и Babesia (Muñoz-Gómez et al., 2019a). К настоящему времени в ранневетвящихся группах, таких как Gregarina и Corallicods, были обнаружены апикомплексные виды с изменчивыми жизненными стратегиями. Однако существование этого, как сообщается, мутуалистического таксона глубоко внутри Apicomplexa, сестры группы высоковирулентных паразитов крови, предполагает, что уникальная биология Nephromycidae может быть ответственна за такой переход к комменсальной или мутуалистической жизненной стратегии.
Cardiosporidium cionae был первоначально описан в 1907 году Ван Гавером и Стефаном, которые правильно определили его как нового споровика-паразита инвазивной оболочника Ciona кишечная . Этот вид больше не упоминался до тех пор, пока он не был обнаружен Сциппой, Чансио и де Винсентиисом в 2000 году, а затем официально переописан Чиансио и др. (2008), спустя целое столетие после его первоначального открытия. Подобно другим гемоспоридиям, таким как Plasmodium , C.cionae обнаруживается в крови хозяина. Он локализуется в сердце и теле перикарда, скоплении отслоившихся клеток, которые накапливаются в течение жизни оболочника внутри перикарда (Evans Anderson and Christiaen, 2016). Ciona кишечная является высокоинвазивным; этот плодовитый вид распространился по всему миру, путешествуя в корпусах и льяльных водах кораблей, и теперь встречается на всех континентах, кроме Антарктиды. В то время как инфекция C. cionae была официально подтверждена только в Неаполитанском заливе, Италия (Ciancio et al. , 2008) и в заливе Наррагансетт, штат Род-Айленд, США, он, вероятно, также имеет широкий ареал. Кроме того, данные ПЭМ при переописании C. cionae выявили бактериальный эндосимбионт (Ciancio et al., 2008).
, 2008) и в заливе Наррагансетт, штат Род-Айленд, США, он, вероятно, также имеет широкий ареал. Кроме того, данные ПЭМ при переописании C. cionae выявили бактериальный эндосимбионт (Ciancio et al., 2008).
Ближайший родственник C. cionae , Nephromyces , был впервые описан примерно в то же время в 1888 году, хотя его необычная нитевидная морфология привела к тому, что до 2010 года его ошибочно классифицировали как хитридиевый гриб (Saffo et al., 2010). Nephromyces обнаружен в семействе оболочников Molgulidae в беспротоковой структуре неизвестной функции, прилегающей к сердцу, известной как почечный мешок.Считается, что он является мутуалистическим из-за почти 100% распространенности инфекции (Saffo et al., 2010) и способен использовать продукты жизнедеятельности, которые оболочка хозяина изолирует в почечном мешке, в качестве источника глицина, пирувата и малата. Пайт и др., 2019). Nephromyces также содержит три отдельные линии бактериальных эндосимбионтов (Paight et al. , 2020). Хотя эндосимбиотические ассоциации обычно встречаются у других простейших, таких как инфузории, диатомовые водоросли и амебы, бактериальный эндосимбиоз у Apicomplexa уникален для этой линии (Nowack and Melkonian, 2010), которая включает только Cardiosporidium и Nephromyces (Muñoz-Gómez et др., 2019а).
, 2020). Хотя эндосимбиотические ассоциации обычно встречаются у других простейших, таких как инфузории, диатомовые водоросли и амебы, бактериальный эндосимбиоз у Apicomplexa уникален для этой линии (Nowack and Melkonian, 2010), которая включает только Cardiosporidium и Nephromyces (Muñoz-Gómez et др., 2019а).
Эндосимбиотические бактерии позволяют эукариотам использовать огромный диапазон сред, в которых они в противном случае не смогли бы обитать. Эндосимбионты охватывают широкий спектр таксонов, от Buchnera эндосимбионтов тлей, которые обеспечивают необходимые витамины и аминокислоты, до хемотрофных бактерий в основе пищевой цепи глубоководных гидротермальных жерл. Разнообразие путей метаболизма прокариот (McCutcheon et al., 2019) обусловливает склонность бактерий колонизировать и использовать необычные места обитания, включая такие экстремальные условия, как радиоактивные отходы (Fredrickson et al., 2004), сильнокислые горячие источники (Marciano-Cabral, 1988) или даже внутри хозяина. У многоклеточных хозяев бактериальные эндосимбионты часто изолированы в специфических структурах или тканях, но у протистов они должны располагаться непосредственно в цитоплазме, что делает эти ассоциации гораздо более тесными (Nowack and Melkonian, 2010).
У многоклеточных хозяев бактериальные эндосимбионты часто изолированы в специфических структурах или тканях, но у протистов они должны располагаться непосредственно в цитоплазме, что делает эти ассоциации гораздо более тесными (Nowack and Melkonian, 2010).
Хотя эти взаимодействия кажутся полезными, эндосимбиоз коренится в конфликте (Keeling and McCutcheon, 2017; McCutcheon et al., 2019). Многие из распространенных эндосимбиотических таксонов, например, относящиеся к отряду Rickettsiales , тесно связаны с патогенами. Rickettsiales , вероятно, является сестринским таксоном современных эукариотических митохондрий (Fitzpatrick et al., 2006), а также содержит Wolbachia , род эндосимбионтов членистоногих и нематод, которые, как известно, инфицируют 25–70% насекомых (Kozek and Rao, 2007). Эндосимбиоз и патогенез тесно связаны из-за механизмов инвазии и персистенции клеток-хозяев (Keeling and McCutcheon, 2017). Однако вторгшиеся бактерии редко видят долгосрочные выгоды от этих взаимодействий. Часто обнаруживается, что геномы эндосимбионтов сильно редуцированы из-за воздействия храповика Мюллера, при котором узкие места в популяции вертикально передающихся эндосимбионтов вызывают накопление вредных мутаций с течением времени (Moran, 1996; Nowack and Melkonian, 2010; McCutcheon and Moran, 2012). .При отсутствии потока генов между популяциями эндосимбионты не могут восстановиться после мутаций и ошибок репликации, которые с большей вероятностью возникают в регионах, богатых G/C, что приводит к характерному смещению A/T (McCutcheon et al., 2019). Чистым воздействием этих сил является создание сильно редуцированных, богатых А/Т геномов, которые конвергентно развились в большинстве вертикально передающихся линий эндосимбионтов (Moran, 1996; Nowack and Melkonian, 2010; McCutcheon and Moran, 2012; Keeling and McCutcheon, 2017; McCutcheon et al., 2019). Хотя эндосимбионта кормят и размещают, он также фактически выведен из строя и навсегда привязан к своему хозяину.
Часто обнаруживается, что геномы эндосимбионтов сильно редуцированы из-за воздействия храповика Мюллера, при котором узкие места в популяции вертикально передающихся эндосимбионтов вызывают накопление вредных мутаций с течением времени (Moran, 1996; Nowack and Melkonian, 2010; McCutcheon and Moran, 2012). .При отсутствии потока генов между популяциями эндосимбионты не могут восстановиться после мутаций и ошибок репликации, которые с большей вероятностью возникают в регионах, богатых G/C, что приводит к характерному смещению A/T (McCutcheon et al., 2019). Чистым воздействием этих сил является создание сильно редуцированных, богатых А/Т геномов, которые конвергентно развились в большинстве вертикально передающихся линий эндосимбионтов (Moran, 1996; Nowack and Melkonian, 2010; McCutcheon and Moran, 2012; Keeling and McCutcheon, 2017; McCutcheon et al., 2019). Хотя эндосимбионта кормят и размещают, он также фактически выведен из строя и навсегда привязан к своему хозяину.
Содержание эндосимбионта также дорого обходится хозяину, а содержание чужеродной клетки вместо ее переваривания или изгнания указывает на то, что эндосимбионт дает значительное преимущество. В рамках более широкого исследования Nephromycidae здесь мы сосредоточимся на характеристике роли бактериального эндосимбионта, о котором сообщалось в C. cionae (Ciancio et al., 2008).Поскольку Cardiosporidium и Nephromyces поддерживали эндосимбионты α-протеобактерий с тех пор, как они дивергировались, мы предполагаем, что эта линия эндосимбионтов должна обеспечивать метаболические функции высокой ценности для своих апикомплексов-хозяев. Поддержание бактериальных эндосимбионтов может снижать зависимость от хозяина и конкуренцию за ресурсы, обеспечивая новые пути биосинтеза, тем самым снижая вирулентность в этой уникальной линии.
В рамках более широкого исследования Nephromycidae здесь мы сосредоточимся на характеристике роли бактериального эндосимбионта, о котором сообщалось в C. cionae (Ciancio et al., 2008).Поскольку Cardiosporidium и Nephromyces поддерживали эндосимбионты α-протеобактерий с тех пор, как они дивергировались, мы предполагаем, что эта линия эндосимбионтов должна обеспечивать метаболические функции высокой ценности для своих апикомплексов-хозяев. Поддержание бактериальных эндосимбионтов может снижать зависимость от хозяина и конкуренцию за ресурсы, обеспечивая новые пути биосинтеза, тем самым снижая вирулентность в этой уникальной линии.
Материалы и методы
Микроскопия
Визуальные скрининги гемолимфы Ciona кишечная были проведены с использованием окраски 5% Гимза/фосфатным буфером с препаратом тонкого мазка, который обычно используется для выявления малярийных инфекций (Moll et al., 2008). Нитевидная стадия жизни была идентифицирована в ходе этих скринингов из-за ее морфологического сходства с Nephromyces . Для подтверждения идентичности три образца, включающие 10–15 представляющих интерес типов клеток, отбирали вручную и промывали с помощью растянутых пастеровских пипеток и фосфатно-солевого буфера. Эти образцы были извлечены, ПЦР амплифицированы с C. cionae специфическими праймерами, и полученный продукт ПЦР секвенирован на платформе Sanger в Центре секвенирования генома Университета Род-Айленда.Для локализации бактериальных эндосимбионтов использовали флуоресцентную гибридизацию in situ (FISH) со специфическими зондами класса 16S рРНК, как показано на рисунках 1E,J. Гибридизацию проводили, как описано Paight et al. (2020).
Для подтверждения идентичности три образца, включающие 10–15 представляющих интерес типов клеток, отбирали вручную и промывали с помощью растянутых пастеровских пипеток и фосфатно-солевого буфера. Эти образцы были извлечены, ПЦР амплифицированы с C. cionae специфическими праймерами, и полученный продукт ПЦР секвенирован на платформе Sanger в Центре секвенирования генома Университета Род-Айленда.Для локализации бактериальных эндосимбионтов использовали флуоресцентную гибридизацию in situ (FISH) со специфическими зондами класса 16S рРНК, как показано на рисунках 1E,J. Гибридизацию проводили, как описано Paight et al. (2020).
. стадия жизни ооцист (D,I) и вертикально переносимые флуоресцентные гибридизация in situ (FISH) меченные бактериальные эндосимбионты внутри ооцист (E,J) .Масштабные линейки являются приблизительными из-за изменения размера изображений. FISH проводили по методу Paight et al. (2020).
Извлечение, секвенирование, сборка и группирование
Материал для транскриптома Cardiosporidium cionae собирали и выделяли из диких оболочников Ciona enteralis , как подробно описано Paight et al. (2019). Градиент плотности сахарозы использовали для выделения C. cionae из оболочечной гемолимфы и для обогащения сильно инфицированных образцов гемолимфы, идентифицированных с помощью микроскопии.Градиент состоял из 20, 25, 30, 35 и 40% сахарозы в фосфатном буфере, загруженного приблизительно 5 мл гемолимфы, и центрифугирован в бакет-роторе при 500×g в течение 30 минут при 4°C (Paight и др., 2019). Затем собирали слои 25 и 30%, осаждали, промывали фосфатно-солевым буфером и хранили при -80°C. РНК экстрагировали из осадка, а в сильно инфицированных образцах использовали набор Zymo Quick-RNA (Zymo Research LLC, Ирвин, Калифорния). Три образца с нефильтрованной гемолимфой, гемолимфой, обогащенной 25%-м слоем, и гемолимфой, обогащенной 30%-м слоем, были отправлены на сухом льду в Университет Мэриленда, Балтиморский институт геномных наук и мультиплексированы на одной дорожке Illumina HiSeq.Эти образцы произвели 92 250 706, 109 023 104 и 110 243 954 чтения (Paight et al., 2019). Они были собраны с помощью Trinity/Trinotate v2.
(2019). Градиент плотности сахарозы использовали для выделения C. cionae из оболочечной гемолимфы и для обогащения сильно инфицированных образцов гемолимфы, идентифицированных с помощью микроскопии.Градиент состоял из 20, 25, 30, 35 и 40% сахарозы в фосфатном буфере, загруженного приблизительно 5 мл гемолимфы, и центрифугирован в бакет-роторе при 500×g в течение 30 минут при 4°C (Paight и др., 2019). Затем собирали слои 25 и 30%, осаждали, промывали фосфатно-солевым буфером и хранили при -80°C. РНК экстрагировали из осадка, а в сильно инфицированных образцах использовали набор Zymo Quick-RNA (Zymo Research LLC, Ирвин, Калифорния). Три образца с нефильтрованной гемолимфой, гемолимфой, обогащенной 25%-м слоем, и гемолимфой, обогащенной 30%-м слоем, были отправлены на сухом льду в Университет Мэриленда, Балтиморский институт геномных наук и мультиплексированы на одной дорожке Illumina HiSeq.Эти образцы произвели 92 250 706, 109 023 104 и 110 243 954 чтения (Paight et al., 2019). Они были собраны с помощью Trinity/Trinotate v2. 4.0 (Haas et al., 2013) и итеративно объединены в бины с помощью OrthoFinder v2.3.3 (Emms and Kelly, 2019) с использованием пользовательской базы данных оболочников, альвеолятов и данных бактериальных эндосимбионтов для удаления загрязнения из хозяин и окружающая среда.
4.0 (Haas et al., 2013) и итеративно объединены в бины с помощью OrthoFinder v2.3.3 (Emms and Kelly, 2019) с использованием пользовательской базы данных оболочников, альвеолятов и данных бактериальных эндосимбионтов для удаления загрязнения из хозяин и окружающая среда.
Для геномного секвенирования штаммов C. intestalis были собраны в Снаг-Харбор в Южном Кингстауне, Род-Айленд (41°23′13.4″N, 71°31′01,5″W) в августе и сентябре 2018 г., следуя тому же протоколу для рассечения и извлечения иглой оболочечной гемолимфы из перикардиального мешка. Градиент плотности сахарозы, описанный выше, также использовали для выделения клеток, инфицированных C. cionae , для геномной ДНК, за исключением того, что в 3 из 4 использованных образцов слой 25% сахарозы был заменен на слой 25% для лучшего захвата C. .cionae инфицированных клеток. В четвертом образце использовался слой 30%. Представляющий интерес слой центрифугировали, собирали, осаждали и промывали, как описано выше, и в Paight et al. (2019). Отфильтрованные образцы использовали отдельно, а не включали в нефильтрованные образцы гемолимфы. Геномную ДНК немедленно экстрагировали с использованием 1% лизирующего буфера SDS, протеиназы К и фенол-хлороформной экстракции с последующим осаждением этанолом в течение ночи при -20°C. Качество и концентрацию образцов оценивали с помощью гель-электрофореза, NanoDrop и Qubit (широкий диапазон), а затем хранили при -20°C.
(2019). Отфильтрованные образцы использовали отдельно, а не включали в нефильтрованные образцы гемолимфы. Геномную ДНК немедленно экстрагировали с использованием 1% лизирующего буфера SDS, протеиназы К и фенол-хлороформной экстракции с последующим осаждением этанолом в течение ночи при -20°C. Качество и концентрацию образцов оценивали с помощью гель-электрофореза, NanoDrop и Qubit (широкий диапазон), а затем хранили при -20°C.
Образцы из четырех отдельных градиентных колонок были индивидуально подготовлены в Центре секвенирования генома Университета Род-Айленда, а полученные библиотеки были запущены на одной дорожке Illumina HiSeq4000 в Университете Мэриленда, Балтиморский институт геномных наук.Эти библиотеки были независимо обрезаны и оценены на качество с помощью Trimmomatic v0.36 и FastQC v0.11.8, а затем объединены и собраны с помощью SPAdes v3.13.0 на сервере OSCAR Университета Брауна (Bolger et al., 2014; «FastQC A Quality Control Tool for Высокопроизводительные данные о последовательностях», без даты; Банкевич и др. , 2012).
, 2012).
Метагеномная сборка SPAdes была выделена путем присвоения таксономии контигам с помощью CAT (von Meijenfeldt et al., 2019). Последовательности Rickettsiales были подтверждены с использованием MetaBAT (Kang et al., 2015), а полученные контиги проверяются на загрязнение и повторно собираются с помощью Geneious v9.1.8 (Geneious, 2017). Дополнительные последовательности апикомплексов были идентифицированы путем сопоставления усеченных и объединенных в ячейки транскриптомных прочтений с полной метагеномной сборкой с использованием Bowtie2 v2.3.5.1 и покрытия контигов, рассчитанного с помощью функции геномеков bedtools v2.26.0 (Quinlan and Hall, 2010; Langmead and Salzberg, 2012). . Полученный файл был отсортирован с помощью R для извлечения контигов с более чем 50% покрытием C.cionae стенограммы. Геномные сборки C. cionae и α-протеобактериальных эндосимбионтов были обрезаны до минимальной длины 1 т.п.н., поскольку маловероятно, чтобы контиги меньшего размера были надежно объединены в бины. Графики сборки генома также визуализировались с помощью Bandage v0.8.1 (Wick et al., 2015), а интересующие кластеры определялись с помощью BLAST. Кластер α-протеобактерий идентифицировали с помощью BLAST, экспортировали и сравнивали с бактериальной сборкой CAT с оценкой средней нуклеотидной идентичности (ANI) (Rodriguez-R and Konstantinidis, 2016).
Графики сборки генома также визуализировались с помощью Bandage v0.8.1 (Wick et al., 2015), а интересующие кластеры определялись с помощью BLAST. Кластер α-протеобактерий идентифицировали с помощью BLAST, экспортировали и сравнивали с бактериальной сборкой CAT с оценкой средней нуклеотидной идентичности (ANI) (Rodriguez-R and Konstantinidis, 2016).
органелларных сборки как для апикопласта, так и для митохондрии были созданы с помощью NOVOPlasty v3.7.2 (Dierckxsens et al., 2016). Последовательности семян для этих сборок были обнаружены с использованием геномов апикопластов Nephromyces (Muñoz-Gómez et al., 2019a) и последовательностей Сэнгера C. cionae гена субъединицы цитохром-С-оксидазы 1 (COX-1), сгенерированных с помощью ПЦР с локальными базами данных BLASTN (Madden, 2003).
Предсказание гена и аннотация
Аннотация генома эндосимбионта α-протеобактерий и C.cionae митохондрий проводили с помощью Prokka v1.14.5 (Seemann, 2014). Близкородственные геномы Rickettsia (на что указывает предварительная филогения 16S) были извлечены из NCBI и использованы для создания пользовательской базы данных для аннотации генома α-протеобактерий (Seemann, 2014; дополнительная таблица S1). Аннотацию апикопласта проводили в Geneious v9.1.8 с использованием пользовательской базы данных аннотаций апикопластов Nephromyces (Muñoz-Gómez et al., 2019a). Области инвертированных повторов были идентифицированы с помощью плагина Repeat Finder.
Аннотацию апикопласта проводили в Geneious v9.1.8 с использованием пользовательской базы данных аннотаций апикопластов Nephromyces (Muñoz-Gómez et al., 2019a). Области инвертированных повторов были идентифицированы с помощью плагина Repeat Finder.
Cardiosporidium cionae геномных контигов были аннотированы с помощью конвейера MAKER v2.31.10 (Holt and Yandell, 2011). Повторы были мягко замаскированы с помощью RepeatMasker v4.0.9 (Smit et al., 2013). Прогнозы Ab initio и параметры обучения видов были сгенерированы как с помощью скрытых марковских моделей WebAugustus (Hoff and Stanke, 2013), так и с помощью скрытых марковских моделей (Korf, 2004). Этот процесс повторялся итеративно, и значения AED, указывающие на соответствие прогноза гена модели, анализировались для обеспечения высокого качества прогнозов.Предсказанные белки обоих организмов были функционально классифицированы с помощью Киотской энциклопедии генов и геномов (KEGG) и NCBI BLASTP v2.7.0+ (Madden, 2003; Kanehisa et al. , 2017). Поиск гомологичных доменов в кодирующих последовательностях проводили с помощью InterProScan (Mitchell et al., 2019). Отдельные интересующие гены подвергали скринингу с использованием баз данных BLAST.
, 2017). Поиск гомологичных доменов в кодирующих последовательностях проводили с помощью InterProScan (Mitchell et al., 2019). Отдельные интересующие гены подвергали скринингу с использованием баз данных BLAST.
Анализ
Полнота генома и транскриптома C. cionae была оценена с помощью BUSCO с использованием эукариотической базы данных (Simão et al., 2015). Гомологи, идентифицированные BUSCO как множественные копии, были вручную проверены с помощью NCBI-BLAST, чтобы подтвердить, что они не представляют загрязнения в готовой сборке. Белки аннотировали с использованием ортологов из EuPathDB, PFAM, Kegg и Interpro. Транскрипты с несколькими предсказанными изоформами в транскриптоме были отфильтрованы и отобраны на основе полноты, оценки Interpro и длины. Полноту и контаминацию генома α-протеобактерий C. cionae оценивали с помощью Атласа микробных геномов (MiGA) и CheckM (Parks et al., 2015; Родригес-Р и др., 2018). Псевдогены-кандидаты были обнаружены с помощью Pseudfinder с параметрами по умолчанию с использованием пользовательской базы данных BLAST белков из 785 полных геномов альфа-протеобактерий, доступных на NCBI (Syberg-Olsen et al.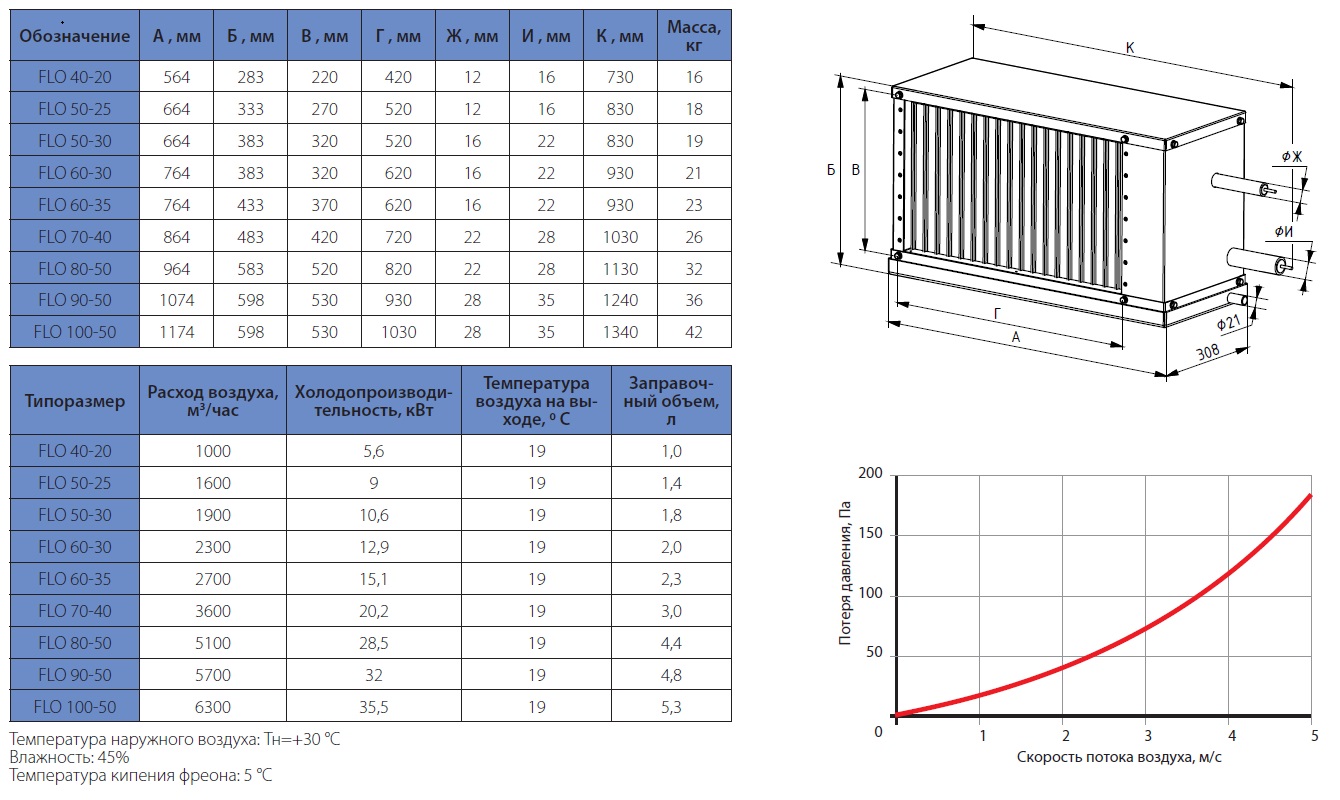 , 2018). Визуальные представления метаболических путей были построены как для C. cionae , так и для α-протеобактерий с использованием функциональных аннотаций из KEGG (Kanehisa et al., 2017). Визуальные представления α-протеобактерий были созданы с помощью Circos (Krzywinski et al., 2009) с данными аннотации от Prokka и функциональными аннотациями от KEGG. Помимо функциональных сравнений с использованием аннотаций KEGG, геном эндосимбионта α-протеобактерий также сравнивали с эндосимбионтом Nephromyces с оценками сходства и содержанием ортологичных генов. Сходство сравнивали с расчетами ANI и средней идентичности аминокислот (AAI) с использованием онлайн-калькулятора ANI и AAI (Rodriguez-R and Konstantinidis, 2016). Сравнение содержания ортологических генов между C.cionae α-протеобактерий и всех эндосимбионтов в системе Nephromyces проводили с помощью OrthoFinder v2.3.3 (Emms and Kelly, 2019). Результирующие перекрытия были рассчитаны с использованием пакета R limma (Ritchie et al.
, 2018). Визуальные представления метаболических путей были построены как для C. cionae , так и для α-протеобактерий с использованием функциональных аннотаций из KEGG (Kanehisa et al., 2017). Визуальные представления α-протеобактерий были созданы с помощью Circos (Krzywinski et al., 2009) с данными аннотации от Prokka и функциональными аннотациями от KEGG. Помимо функциональных сравнений с использованием аннотаций KEGG, геном эндосимбионта α-протеобактерий также сравнивали с эндосимбионтом Nephromyces с оценками сходства и содержанием ортологичных генов. Сходство сравнивали с расчетами ANI и средней идентичности аминокислот (AAI) с использованием онлайн-калькулятора ANI и AAI (Rodriguez-R and Konstantinidis, 2016). Сравнение содержания ортологических генов между C.cionae α-протеобактерий и всех эндосимбионтов в системе Nephromyces проводили с помощью OrthoFinder v2.3.3 (Emms and Kelly, 2019). Результирующие перекрытия были рассчитаны с использованием пакета R limma (Ritchie et al. , 2015), а окончательная цифра сгенерирована с помощью диаграммы Венна (Chen and Boutros, 2011), также в R. Функциональное перекрытие генов было основано на аннотациях KEGG и сгенерировано с использованием те же пакеты R.
, 2015), а окончательная цифра сгенерирована с помощью диаграммы Венна (Chen and Boutros, 2011), также в R. Функциональное перекрытие генов было основано на аннотациях KEGG и сгенерировано с использованием те же пакеты R.
Филогенетика
Бактериальные филогении были построены с использованием предсказанной таксономии от MiGA, которая отнесла эндосимбионт к классу альфапротеобактерий со значением p , равным 0.25 (Паркс и др., 2015; Родригес-Р и др., 2018). Чтобы подтвердить этот результат, все полные образцы бактериального протеома, принадлежащие к этому классу, были извлечены из базы данных NCBI (всего 712). Поиск этих данных проводился с использованием набора единичных копий генов HMM α-протеобактерий, состоящего из 117 белков, выровненных, и дерева, построенного с использованием рабочего процесса GToTree (Lee, 2019; «Accelerated Profile HMM Searches» nd; Edgar, 2004; Hyatt et al. , 2010 г., Прайс и др., 2010 г., «TaxonKit — инструментарий таксономии NCBI, n. д.; Капелла-Гутьеррес и др., 2009).
д.; Капелла-Гутьеррес и др., 2009).
Кодируемые апикопластами гены C. cionae были добавлены к набору данных, использованному Muñoz-Gómez et al. (2019a), чтобы подтвердить монофилию с Nephromyces , ранее указанную с помощью деревьев генов COI и 18S. Гомологи белка были идентифицированы с использованием локального поиска BLAST-P и объединены с существующим набором данных. Эти последовательности были выровнены с MAFFT v7, обрезаны в Geneious и объединены (Madden, 2003; Katoh and Standley, 2013). Филогенез вида был выведен с максимальной вероятностью с использованием IQ-TREE (v1.6) и модель LG+G. Статистическая поддержка в филиалах оценивалась с использованием сверхбыстрой начальной загрузки (1000) и aLRT (1000) (Nguyen et al., 2015).
Параметры
Конкретные сценарии и настройки, используемые для биоинформатического анализа C. cionae и его эндосимбионта, были размещены в общедоступном репозитории GitHub (github.com/liz-hunter/cardio_project).
Результаты
Cardiosporidium cionae Геномное секвенирование пула C.cionae дали в общей сложности 320 000 000 парных прочтений. После обрезки и сборки в результате получился 656 251 контиг, 176 701 из которых были больше 1 т.п.о. Биннинг с помощью CAT привел к тому, что 3641 контиг был отнесен к суперфилуму Alveolata. Контиги, отнесенные к Dinophyceae и Ciliophora, были удалены, оставив 2754 контига, и 1790 из этих контигов были больше 1 т.п.о. Биннинг на основе покрытия с помощью RNA-seq добавил дополнительные 935 контигов, 423 из которых были уникальными и размером более 1 т.п.н.Дальнейшее ручное курирование с помощью OrthoFinder устранило 7 дополнительных контигов. В результате всего 2206 контигов были отнесены к C. cionae. Из оставшихся 174 496 контигов 221 относили к порядку Rickettsiales, а 147 793 контига относили к классу Ascidiacea (оболочечные). Остальные контиги, которые были классифицированы CAT, были либо не классифицированы, либо идентифицированы как загрязнение различными морскими организмами, особенно бактериями и другими простейшими. Хотя мы не можем исключить морскую воду как источник загрязнения, это, вероятно, связано с физиологией оболочников, таких как Ciona кишечная , которые являются фильтраторами с открытой системой кровообращения.Сборка генома C. cionae составляет в целом 57 Мб, с N50 54,04 т.п.н. и содержанием G/C 34,4% (таблица 1). Это меньше, чем у некоторых апикомплексных геномов, таких как кокцидий Toxoplasma gondi (80 Мб), но значительно больше, чем у гемоспоридий Plasmodium falciparum (22,9 Мб) и сильно редуцированных Cryptosporidium parvum (9 Мб) (Sibley and Boothroyd, 1999). ; Абрахамсен и др., 2004). Предсказание генов привело к 4674 белкам (таблица 1). Сборка объединенного транскриптома дала в общей сложности 15 077 белков, относящихся к C.cionae , включая все изоформы. После фильтрации для удаления избыточности этот набор данных был сокращен до 6733 уникальных белков.
Хотя мы не можем исключить морскую воду как источник загрязнения, это, вероятно, связано с физиологией оболочников, таких как Ciona кишечная , которые являются фильтраторами с открытой системой кровообращения.Сборка генома C. cionae составляет в целом 57 Мб, с N50 54,04 т.п.н. и содержанием G/C 34,4% (таблица 1). Это меньше, чем у некоторых апикомплексных геномов, таких как кокцидий Toxoplasma gondi (80 Мб), но значительно больше, чем у гемоспоридий Plasmodium falciparum (22,9 Мб) и сильно редуцированных Cryptosporidium parvum (9 Мб) (Sibley and Boothroyd, 1999). ; Абрахамсен и др., 2004). Предсказание генов привело к 4674 белкам (таблица 1). Сборка объединенного транскриптома дала в общей сложности 15 077 белков, относящихся к C.cionae , включая все изоформы. После фильтрации для удаления избыточности этот набор данных был сокращен до 6733 уникальных белков.
Таблица 1. Представлены статистические данные для геномных и транскриптомных наборов данных.
Окончательная сборка генома C. cionae , по оценкам BUSCO, завершена на 63,7%, при этом 13,9% дублируются BUSCO. Транскриптом немного более полный: по оценке BUSCO, 68,3% полных ортологов и 12,5% частичных (Paight et al., 2019). Когда изоформы были отфильтрованы для аннотации, это значение полноты немного снизилось до 60,0% с дублированием 4,6% (таблица 1). Несмотря на это, сборка C. cionae содержит гены всех ожидаемых основных путей биосинтеза гематозоя (дополнительная фигура S3). Cardiosporidium имеет набор основных метаболических путей, включая полные или почти полные функциональные прогнозы для гликолиза, глюконеогенеза, окисления пирувата, пентозофосфатного цикла и цикла лимонной кислоты (рисунок 2 и дополнительная таблица S2).Он также кодирует несколько неожиданных путей, включая весь путь биосинтеза de novo IMP. Cardiosporidium содержит гены биосинтеза и удлинения жирных кислот в эндоплазматическом ретикулуме, а также деградации с образованием ацетил-КоА (рисунок 2 и дополнительная таблица S2).
Рисунок 2. Метаболизм Cardiosporidium cionae . Сплошные цвета обозначают гомологи геномных белков, точки показывают, где гомологи были обнаружены только в данных транскриптома, а жирным шрифтом обозначены пути бактериального эндосимбионта.Эта цифра соответствует информации о геноме и транскриптоме в дополнительной таблице 2.
Cardiosporidium также кодирует большую часть пути биосинтеза триацилглицерина и частичные пути синтеза холестерина и кетоновых тел. У него полностью отсутствуют какие-либо свидетельства наличия генов биосинтеза восьми из двадцати одной аминокислоты, но он кодирует пути превращения аминокислот, которых нет у других гематозоидов. К ним относятся превращение фенилаланина в тирозин и гомоцистеина в метионин (рисунок 2 и дополнительная таблица S2).Кроме того, C. cionae способен генерировать серин из нескольких источников (глицерат-3P и глицин), а также расщеплять его до пирувата. Геномные данные, которые мы восстановили, кодируют только частичные пути синтеза рибофлавина и гема, а также отсутствуют гены синтеза биотина, тиамина, убихинона и кобаламина. Однако мы идентифицировали пути биосинтеза изопреноидов С5 и С10-20. Этот геном также подтверждает наличие пути деградации пуринов, ранее идентифицированного в транскриптоме Nephromyces (Paight et al., 2019).
Однако мы идентифицировали пути биосинтеза изопреноидов С5 и С10-20. Этот геном также подтверждает наличие пути деградации пуринов, ранее идентифицированного в транскриптоме Nephromyces (Paight et al., 2019).
Визуальные скрининги с окрашиванием тонкого мазка по Гимзе показывают, что C. cionae сохраняет очень низкую плотность внутри хозяина. Эти скрининговые микроскопии дополнительно выявили наличие крупной внеклеточной нитевидной стадии жизни, аналогичной нитевидной стадии жизни Nephromyces (рис. 1). Выделение отдельных клеток, экстракция и ПЦР подтвердили, что эти типы клеток действительно являются жизненной стадией C. cionae .
Филогенетический анализ кодируемых апикопластом белков подтвердил монофилию Nephromyces и C.cionae (рис. 3). Этот анализ отличается от результатов размещения для Nephromyces , опубликованных Muñoz-Gómez et al. (2019a) из-за максимальных данных и исключения таксонов раннего ветвления. Эта выборка таксона привела к тому, что Nephromycidae разветвились за пределы Hematozoa. Апикопласт C. cionae структурно очень похож на таковой Nephromyces с точки зрения содержания генов, размера и организации (дополнительный рисунок S1).
Эта выборка таксона привела к тому, что Nephromycidae разветвились за пределы Hematozoa. Апикопласт C. cionae структурно очень похож на таковой Nephromyces с точки зрения содержания генов, размера и организации (дополнительный рисунок S1).
Рис. 3. Филогения апикопластов, созданная с использованием модифицированного набора данных, предоставленного Muñoz-Gómez et al. (2019a), демонстрирующие монофилию C. cionae и Nephromyces . Полный циркуляризованный апикопласт C. cionae , извлеченный из набора геномных данных, и апикопластов Nephromyces sp. 654, показаны синим цветом. Разрешенная топология отличается от Muñoz-Gómez et al. (2019a) из-за исключения грегарин из нашего набора данных, который включает Nephromycida (синий), Piroplasmida (красный), Coccidia (зеленый) и апикомонады (желтый) в качестве внешней группы.Статистическая поддержка была оценена с использованием aLRT (1000) и сверхбыстрой начальной загрузки (1000) (Nguyen et al. , 2015), и эти значения показаны в этом порядке на узлах.
, 2015), и эти значения показаны в этом порядке на узлах.
α-протеобактерии
Из контигов, отнесенных к Rickettsiales, 45 были больше 1 т.п.о. В результате повторной сборки был получен 31 контиг, а ручная обработка привела к окончательным 29 контигам. Окончательная сборка α-эндосимбионта составляет всего 1,05 млн. п.о., с N50 250,39 т.п.н. и содержанием G/C 29,1%. Предсказание и аннотация генов привели к 906 белкам (таблица 1).Было показано, что кластер Bandage имеет ANI (двусторонний ANI) 99,95% (SD, 81%) на основе 4878 фрагментов по сравнению с бинарной сборкой CAT. Это обеспечило независимую проверку биннинга сборки бактериального генома. По оценкам, сборка α-эндосимбионта завершена на 91,9% при 1,8%-м загрязнении MiGA и на 95,5% при 2,1%-м загрязнении CheckM.
Характерен для бактериальных эндосимбионтов, имеет низкое содержание G/C и высокую плотность кодирования (рис. 4). Прокка предсказал, что этот организм кодирует всего 906 генов (таблица 1), 37 из которых были идентифицированы псевдогенами-кандидатами псевдогенами. Большинство этих псевдогенов происходят от генов с неизвестной функцией, в то время как некоторые происходят от генов, кодирующих пермеазу, транспозазу, тиоэстеразу, фосфодиэстеразу и множественные трансферазы. Pseudfinder также присоединил 13 ORF, оставив только 865 предсказанных генов. С таким небольшим количеством генов неудивительно, что у этой α-протеобактерии небольшое количество полных метаболических путей (рис. 2, 4 и дополнительная рис. S3). Геном немного меньше, чем у близкородственных альфапротеобактериальных эндосимбионтов, таких как Candidatus Phycorickettsia trachydisci sp.ноябрь (1,4 МБ), Orientia tsutsugamushi (2 МБ) и другие связанные с протистами линии Rickettsiales (1,4–1,7 МБ) (Nakayama et al., 2010; Юрченко и др., 2018; Muñoz-Gómez et al., 2019b) . Геномы α-протеобактерий как C. cionae , так и Nephromyces кодируют пути биосинтеза жирных кислот, пиримидинов, липоевой кислоты, гема, глутамина, лизина, убихинона и цикла лимонной кислоты.
Большинство этих псевдогенов происходят от генов с неизвестной функцией, в то время как некоторые происходят от генов, кодирующих пермеазу, транспозазу, тиоэстеразу, фосфодиэстеразу и множественные трансферазы. Pseudfinder также присоединил 13 ORF, оставив только 865 предсказанных генов. С таким небольшим количеством генов неудивительно, что у этой α-протеобактерии небольшое количество полных метаболических путей (рис. 2, 4 и дополнительная рис. S3). Геном немного меньше, чем у близкородственных альфапротеобактериальных эндосимбионтов, таких как Candidatus Phycorickettsia trachydisci sp.ноябрь (1,4 МБ), Orientia tsutsugamushi (2 МБ) и другие связанные с протистами линии Rickettsiales (1,4–1,7 МБ) (Nakayama et al., 2010; Юрченко и др., 2018; Muñoz-Gómez et al., 2019b) . Геномы α-протеобактерий как C. cionae , так и Nephromyces кодируют пути биосинтеза жирных кислот, пиримидинов, липоевой кислоты, гема, глутамина, лизина, убихинона и цикла лимонной кислоты. Только α-протеобактерий C. cionae поддерживают гены биосинтеза аспарагина, гликолиза и пентозофосфатного пути (дополнительная фигура S2), в то время как только α-протеобактерий Nephromyces могут завершить биосинтез глутаминовой кислоты.
Только α-протеобактерий C. cionae поддерживают гены биосинтеза аспарагина, гликолиза и пентозофосфатного пути (дополнительная фигура S2), в то время как только α-протеобактерий Nephromyces могут завершить биосинтез глутаминовой кислоты.
Рисунок 4. Размер, распределение контигов, плотность кодирования и аннотации основных функциональных категорий генов в геноме эндосимбионтов α-протеобактерий.
Когда α-протеобактерии в Cardiosporidium и Nephromyces сравнили на предмет сходства, результаты показали, что эти таксоны слишком расходятся, чтобы их можно было сравнивать с ANI, и вместо этого их сравнивали со средним AAI. Двусторонний анализ AAI 656 белков показал 47.61% (стандартное отклонение: 12,51%) сходства между этими геномами, что согласуется с филогенетическим анализом, указывающим на значительную эволюционную дистанцию между этими двумя таксонами. Эта мультигенная филогения α-эндосимбионтов согласуется с предварительными генными деревьями 16S, которые помещают эти виды в порядок Rickettsiales. Они принадлежат к семейству Rickettsiaceae и являются сестрами рода Rickettsia (рис. 5).
Они принадлежат к семейству Rickettsiaceae и являются сестрами рода Rickettsia (рис. 5).
Рисунок 5. Филогенез альфапротеобактерий , созданный с помощью конвейера GToTree (117 соединенных генов), включая все секвенированные альфапротеобактерии, опубликованные на NCBI, и оба C.cionae и Nephromyces α-эндосимбионты (красный). Поддержка начальной загрузки отображается в виде десятичного значения на узлах.
Сравнениеортологов между α-эндосимбионтами показывает, что эти таксоны разделяют большинство своих основных функций, но система Cardiosporidium α-эндосимбионтов поддерживает более уникальные гены. Этот таксон также имеет большее ортологическое и функциональное перекрытие с двумя дополнительными эндосимбионтами, присутствующими в системе Nephromyces : бетапротеобактериями и Bacteroides (дополнительный рисунок S2).
Доступность данных
Все данные, связанные с этим проектом, хранятся в GenBank под номером BioProject PRJNA664590. Проект полного генома Cardiosporidium cionae был депонирован под регистрационным номером JADAQX000000000, геном альфапротеобактериального эндосимбионта депонирован под регистрационным номером JADAQY000000000, а транскриптом депонирован под регистрационным номером GIVE00000000. В этом документе описываются версии JADAQX010000000, JADAQY010000000 и GIVE01000000.
Проект полного генома Cardiosporidium cionae был депонирован под регистрационным номером JADAQX000000000, геном альфапротеобактериального эндосимбионта депонирован под регистрационным номером JADAQY000000000, а транскриптом депонирован под регистрационным номером GIVE00000000. В этом документе описываются версии JADAQX010000000, JADAQY010000000 и GIVE01000000.
Обсуждение
Метаболически C. cionae сходен с другими секвенированными гематозоидами. Однако он также кодирует некоторые необычные пути. Cardiosporidium cionae , как и Nephromyces , кодирует путь биосинтеза пурина de novo (рис. 2 и дополнительная таблица S2), который был утерян во всех других секвенированных апикомплексанах (Janouskovec and Keeling, 2016). Эти гены разрешаются с Nephromyces, Vitrella brassicaformis и динофлагеллятами, такими как Crypthecodinium cohnii , в филогенетическом анализе, демонстрируя, что это не было недавним событием горизонтального переноса генов (Paight et al. , 2020). Вместо этого эти данные указывают на то, что оба рода внутри Nephromycidae сохранили наследственный путь, обнаруженный у свободноживущих Chromerid, а гены биосинтеза пуринов были потеряны независимо во всех других линиях апикомплексов. Биосинтез пуринов De novo в C. cionae и Nephromyces снижает зависимость от предварительно сформированных пуриновых метаболитов у их соответствующих хозяев, потенциально обеспечивая сохранение внеклеточных стадий жизни в обеих этих линиях. Nephromyces и C. cionae также способны расщеплять пурины (Paight et al., 2019), и мы подозреваем, что этот аспект их метаболизма связан с физиологией оболочников, которые не способны метаболизировать мочевую кислоту, пуриновую кислоту. отходы. Несмотря на то, что оболочники не способны расщеплять мочевую кислоту, они необъяснимым образом накапливают ее (Nolfi, 1970; Lambert et al., 1998).
, 2020). Вместо этого эти данные указывают на то, что оба рода внутри Nephromycidae сохранили наследственный путь, обнаруженный у свободноживущих Chromerid, а гены биосинтеза пуринов были потеряны независимо во всех других линиях апикомплексов. Биосинтез пуринов De novo в C. cionae и Nephromyces снижает зависимость от предварительно сформированных пуриновых метаболитов у их соответствующих хозяев, потенциально обеспечивая сохранение внеклеточных стадий жизни в обеих этих линиях. Nephromyces и C. cionae также способны расщеплять пурины (Paight et al., 2019), и мы подозреваем, что этот аспект их метаболизма связан с физиологией оболочников, которые не способны метаболизировать мочевую кислоту, пуриновую кислоту. отходы. Несмотря на то, что оболочники не способны расщеплять мочевую кислоту, они необъяснимым образом накапливают ее (Nolfi, 1970; Lambert et al., 1998).
Принимая во внимание, что полные пути пентозофосфатного цикла, цикла лимонной кислоты и глюконеогенеза отражают другие кроветворные (рисунок 2 и дополнительная таблица S2), C. cionae также кодирует несколько генов, которые предполагают, что он способен производить глиоксилат. Пэйт и др. (2019) сообщили о транскриптах ряда пероксисомальных белков как у C. cionae , так и у Nephromyces , и предсказали новый метаболический путь. Несмотря на их многочисленные метаболические сходства, C. cionae и Nephromyces , по-видимому, имеют разные пути центрального углеродного метаболизма (в частности, цикл лимонной кислоты) и являются частью тесно связанного глиоксилатного цикла.Как Nephromyces , так и C. cionae обладают одинаково высоко выраженным циклом деградации пуринов, который превращает уреидогликолят в глиоксилат с использованием новой амидогидролазы и генерирует глицин и серин. Однако только Nephromyces могут подавать глиоксилат обратно в цикл лимонной кислоты с помощью малатсинтазы (Paight et al., 2019). Этот путь является продуктом необычной среды почечного мешка, где обитает Nephromyces , который содержит большое количество мочевой кислоты, секвестрированной оболочником хозяина.
cionae также кодирует несколько генов, которые предполагают, что он способен производить глиоксилат. Пэйт и др. (2019) сообщили о транскриптах ряда пероксисомальных белков как у C. cionae , так и у Nephromyces , и предсказали новый метаболический путь. Несмотря на их многочисленные метаболические сходства, C. cionae и Nephromyces , по-видимому, имеют разные пути центрального углеродного метаболизма (в частности, цикл лимонной кислоты) и являются частью тесно связанного глиоксилатного цикла.Как Nephromyces , так и C. cionae обладают одинаково высоко выраженным циклом деградации пуринов, который превращает уреидогликолят в глиоксилат с использованием новой амидогидролазы и генерирует глицин и серин. Однако только Nephromyces могут подавать глиоксилат обратно в цикл лимонной кислоты с помощью малатсинтазы (Paight et al., 2019). Этот путь является продуктом необычной среды почечного мешка, где обитает Nephromyces , который содержит большое количество мочевой кислоты, секвестрированной оболочником хозяина. У C. cionae малатсинтаза явно отсутствует как в геноме, так и в транскриптоме, что указывает на то, что круговорот углерода в этих близкородственных организмах, вероятно, отличается и, возможно, является одним из различий, объясняющих несоответствие вирулентности между C. cionae . и Нефромицеты. Однако список различий, включающий также виды хозяев и локализацию органелл, относительно невелик. Хотя было известно, что эти таксоны имеют сходные черты жизненного цикла (Ciancio et al., 2008; Saffo et al., 2010), эти данные позволяют предположить, что их морфология и метаболизм также очень похожи.
У C. cionae малатсинтаза явно отсутствует как в геноме, так и в транскриптоме, что указывает на то, что круговорот углерода в этих близкородственных организмах, вероятно, отличается и, возможно, является одним из различий, объясняющих несоответствие вирулентности между C. cionae . и Нефромицеты. Однако список различий, включающий также виды хозяев и локализацию органелл, относительно невелик. Хотя было известно, что эти таксоны имеют сходные черты жизненного цикла (Ciancio et al., 2008; Saffo et al., 2010), эти данные позволяют предположить, что их морфология и метаболизм также очень похожи.
Cardiosporidium cionae и Nephromyces (Nephromycidae) разветвляются в составе Hematozoa, группы облигатных паразитических внутриклеточных апикомплексов (Muñoz-Gómez et al., 2019a; Mathur et al., 2019). Все ранее описанные представители Hematozoa и сестринского таксона Coccidia являются внутриклеточными и облигатно паразитирующими. Несмотря на свое филогенетическое положение внутри облигатно внутриклеточной клады, представители Nephromycidae имеют большие нитевидные внеклеточные стадии жизни (рис. 1). Nephromyces является полностью внеклеточным (Saffo and Nelson, 1983), тогда как C. cionae имеет как внутриклеточную, так и внеклеточную стадии жизни. Хотя морфологически они сходны с более базальными грегариновыми апикомплексами (Rueckert et al., 2015), эти группы филогенетически далеки. Nephromycidae произошли от внутриклеточных предков и перешли во внеклеточную среду. У Nephromyces этот переход завершен, тогда как у C. cionae есть как внутриклеточная, так и внеклеточная стадии жизни.Мы полагаем, что внеклеточность в этой группе связана с другой необычной характеристикой: сохранением бактериальных эндосимбионтов как у C. cionae , так и у Nephromyces.
1). Nephromyces является полностью внеклеточным (Saffo and Nelson, 1983), тогда как C. cionae имеет как внутриклеточную, так и внеклеточную стадии жизни. Хотя морфологически они сходны с более базальными грегариновыми апикомплексами (Rueckert et al., 2015), эти группы филогенетически далеки. Nephromycidae произошли от внутриклеточных предков и перешли во внеклеточную среду. У Nephromyces этот переход завершен, тогда как у C. cionae есть как внутриклеточная, так и внеклеточная стадии жизни.Мы полагаем, что внеклеточность в этой группе связана с другой необычной характеристикой: сохранением бактериальных эндосимбионтов как у C. cionae , так и у Nephromyces.
Сохранение монофилетических α-эндосимбионтов в обеих линиях C. cionae и Nephromyces указывает на то, что этот эндосимбионт обеспечивает что-то жизненно важное для системы. Однако, на первый взгляд, эти эндосимбионты вносят очень небольшой вклад в апикомплексы своего хозяина. Как и его аналог в Nephromyces , C. cionae α-эндосимбионт содержит лишь несколько путей биосинтеза (рис. 4). В целом, α-эндосимбионт в C. cionae действительно содержит больше уникальных ортологов и функциональных генов по сравнению с его аналогом в системе Nephromyces (дополнительная фигура S2). В первую очередь эти уникальные гены связаны с энергетическим метаболизмом (рисунок 4 и дополнительный рисунок S2), и их присутствие, вероятно, является результатом повышенного эволюционного давления для поддержания критических генов в системе с одним эндосимбионтом по сравнению с тремя типами эндосимбионтов. присутствует в Nephromyces сообществах.Кодируемые α-эндосимбионтом пути для энергетического и углеродного цикла, хотя, возможно, выгодны для C. cionae , вероятно, не являются критическими вкладами, поскольку они могут быть завершены апикомплексаном, независимо от эндосимбионта (дополнительная фигура S3). Поддержание эндосимбионта обходится дорого, и маловероятно, что он будет сохранен для избыточной функции (McCutcheon and Moran, 2012).
Как и его аналог в Nephromyces , C. cionae α-эндосимбионт содержит лишь несколько путей биосинтеза (рис. 4). В целом, α-эндосимбионт в C. cionae действительно содержит больше уникальных ортологов и функциональных генов по сравнению с его аналогом в системе Nephromyces (дополнительная фигура S2). В первую очередь эти уникальные гены связаны с энергетическим метаболизмом (рисунок 4 и дополнительный рисунок S2), и их присутствие, вероятно, является результатом повышенного эволюционного давления для поддержания критических генов в системе с одним эндосимбионтом по сравнению с тремя типами эндосимбионтов. присутствует в Nephromyces сообществах.Кодируемые α-эндосимбионтом пути для энергетического и углеродного цикла, хотя, возможно, выгодны для C. cionae , вероятно, не являются критическими вкладами, поскольку они могут быть завершены апикомплексаном, независимо от эндосимбионта (дополнительная фигура S3). Поддержание эндосимбионта обходится дорого, и маловероятно, что он будет сохранен для избыточной функции (McCutcheon and Moran, 2012).
Несколько путей сохранились в обеих линиях α-эндосимбионтов и также отсутствуют у апикомплексов хозяина.Единственными критически важными функциями, которые не могут быть заменены метаболизмом апикомплексана, являются биосинтез лизина и биосинтез липоевой кислоты. Лизин является незаменимой аминокислотой и играет важную роль в биосинтезе белка. Лизин является важным компонентом среды для роста P. falciparum и, по прогнозам, вымывается из хозяина T. gondii (Schuster, 2002; Тимошенко и др., 2015). Биосинтез лизина также отсутствует в геноме и транскриптоме Nephromyces (Paight et al., 2020). Как и Nephromyces, T. gondii и P. falciparum , наши данные показывают, что C. cionae не может синтезировать собственный лизин и зависит от удаления хозяином. Хотя мы не можем исключить возможность того, что C. cionae кодирует биосинтез лизина с неполным геномом, судя по геномам других апикомплексов, биосинтез лизина, вероятно, отсутствует у Nephromcyidae. Лизин также необходим для оболочника хозяина, Ciona кишечная (Kanehisa et al., 2017), и оба организма, нуждающиеся в источниках лизина из окружающей среды, ставят их в постоянную конкуренцию за этот ресурс. Cardiosporidium cionae , по-видимому, обошел этот конфликт, сохранив бактериальный эндосимбионт, который содержит путь биосинтеза de novo лизина. Вместо того, чтобы конкурировать с оболочником хозяина за лизин, C. cionae культивирует внутриклеточный источник незаменимой аминокислоты, снижая зависимость хозяина и потенциально вирулентность.
Лизин также необходим для оболочника хозяина, Ciona кишечная (Kanehisa et al., 2017), и оба организма, нуждающиеся в источниках лизина из окружающей среды, ставят их в постоянную конкуренцию за этот ресурс. Cardiosporidium cionae , по-видимому, обошел этот конфликт, сохранив бактериальный эндосимбионт, который содержит путь биосинтеза de novo лизина. Вместо того, чтобы конкурировать с оболочником хозяина за лизин, C. cionae культивирует внутриклеточный источник незаменимой аминокислоты, снижая зависимость хозяина и потенциально вирулентность.
Липоевая кислота представляет собой ароматическое соединение серы, которое является важным кофактором для ряда жизненно важных метаболических функций. К ним относятся цикл лимонной кислоты и комплексы альфа-кетодегидрогеназы, такие как комплекс пируватдегидрогеназы и система превращения глицина. У эукариот липоевая кислота локализована исключительно в митохондриях и пластидах. Apicomplexans локализуют биосинтез липоевой кислоты в апикопласте, утратив митохондриальный путь после приобретения пластиды (Crawford et al. , 2006). Вместо этого для производства липоевой кислоты, необходимой для цикла лимонной кислоты и системы превращения глицина в митохондриях, используется альтернативный путь очистки, и оба пути очистки и биосинтеза считаются важными (Günther et al., 2005). Функциональные исследования показали, что когда пути биосинтеза липоевой кислоты отключены, P. falciparum будет компенсировать это за счет удаления большего количества липоевой кислоты из хозяина и доставки ее в апикопласт (Günther et al., 2007). Сходным образом, рост T. gondii ингибируется средой с дефицитом липоата, что позволяет предположить, что необходима чистка (Crawford et al., 2006). Метаболическое моделирование также показывает, что даже апикомплексы, которые поддерживают этот путь, нуждаются в дополнительной липоевой кислоте от своих организмов-хозяев (Blume and Seeber, 2018). Хотя липоевая кислота вырабатывается оболочниками хозяина, мы предполагаем, что ее доступность для внеклеточного организма ограничена, поскольку она вырабатывается и используется в митохондриях.
, 2006). Вместо этого для производства липоевой кислоты, необходимой для цикла лимонной кислоты и системы превращения глицина в митохондриях, используется альтернативный путь очистки, и оба пути очистки и биосинтеза считаются важными (Günther et al., 2005). Функциональные исследования показали, что когда пути биосинтеза липоевой кислоты отключены, P. falciparum будет компенсировать это за счет удаления большего количества липоевой кислоты из хозяина и доставки ее в апикопласт (Günther et al., 2007). Сходным образом, рост T. gondii ингибируется средой с дефицитом липоата, что позволяет предположить, что необходима чистка (Crawford et al., 2006). Метаболическое моделирование также показывает, что даже апикомплексы, которые поддерживают этот путь, нуждаются в дополнительной липоевой кислоте от своих организмов-хозяев (Blume and Seeber, 2018). Хотя липоевая кислота вырабатывается оболочниками хозяина, мы предполагаем, что ее доступность для внеклеточного организма ограничена, поскольку она вырабатывается и используется в митохондриях. Это, вероятно, означает, что C. cionae зависит от этого α-эндосимбионта для производства ключевых соединений, таких как липоевая кислота, для сохранения стабильной стадии внеклеточной жизни. Таким образом, сохранение α-эндосимбионта в качестве внутреннего источника кофактора дополнительно снижает конкуренцию за ресурсы между C. cionae и его хозяином.
Это, вероятно, означает, что C. cionae зависит от этого α-эндосимбионта для производства ключевых соединений, таких как липоевая кислота, для сохранения стабильной стадии внеклеточной жизни. Таким образом, сохранение α-эндосимбионта в качестве внутреннего источника кофактора дополнительно снижает конкуренцию за ресурсы между C. cionae и его хозяином.
Nephromycidae произошли от клады облигатно паразитирующих внутриклеточных апикомплексов и перешли к преимущественно внеклеточному образу жизни.Мы предполагаем, что, получив бактериальные эндосимбионты, эти апикомплексы приобрели метаболические способности, которые сделали возможным этот переход. Хотя Nephromyces имеет общую линию α-эндосимбионтов с C. cionae (рис. 5), у него также есть эндосимбионты бетапротеобактерий и Bacteroides. С этим бактериальным таксономическим разнообразием связано метаболическое разнообразие, и хотя α-эндосимбионт в C. cionae имеет больше уникальных функциональных белков и ортологов, чем его аналог, это ничтожно мало по сравнению с количеством уникальных белков и ортологов, внесенных двумя дополнительными присутствующими таксонами. в системе Nephromyces (дополнительный рисунок S2).Мы полагаем, что единственный эндосимбионт в C. cionae обеспечивает специальный источник основных метаболитов лизина и липоевой кислоты, что, вероятно, снижает конкуренцию с хозяином по сравнению с его гематозойными родственниками и делает возможными внеклеточные стадии жизни. Таким образом, Cardiosporidium cionae представляет собой потенциальное промежуточное звено в переходе к мутуализму, который был описан у Nephromyces (Saffo et al., 2010).
в системе Nephromyces (дополнительный рисунок S2).Мы полагаем, что единственный эндосимбионт в C. cionae обеспечивает специальный источник основных метаболитов лизина и липоевой кислоты, что, вероятно, снижает конкуренцию с хозяином по сравнению с его гематозойными родственниками и делает возможными внеклеточные стадии жизни. Таким образом, Cardiosporidium cionae представляет собой потенциальное промежуточное звено в переходе к мутуализму, который был описан у Nephromyces (Saffo et al., 2010).
Заявление о доступности данных
Авторы признают, что данные, представленные в этом исследовании, должны быть депонированы и сделаны общедоступными в приемлемом хранилище до публикации.Frontiers не может принять рукопись, которая не соответствует нашей политике открытых данных.
Вклад авторов
EH создал и проанализировал набор геномных данных C. cionae и бактериальных эндосимбионтов и провел сравнительный анализ. CP собрала транскриптом C. cionae и бактериальные геномы Nephromyces . CL предоставила финансирование, экспериментальную основу и опыт. Все авторы внесли свой вклад в статью и одобрили представленную версию.
CP собрала транскриптом C. cionae и бактериальные геномы Nephromyces . CL предоставила финансирование, экспериментальную основу и опыт. Все авторы внесли свой вклад в статью и одобрили представленную версию.
Финансирование
Эта работа была поддержана Национальным научным фондом (№ 1541510) и Национальным институтом здравоохранения (R03AI124092).
Конфликт интересов
Авторы заявляют, что исследование проводилось при отсутствии каких-либо коммерческих или финансовых отношений, которые могли бы быть истолкованы как потенциальный конфликт интересов.
Благодарности
Этот материал основан на работе, проведенной в исследовательском центре NSF EPSCoR в Род-Айленде, Центре геномики и секвенирования (GSC), при частичной поддержке Соглашения о сотрудничестве Национального научного фонда EPSCoR #OIA-1655221.В частности, мы хотели бы поблагодарить Джанет Атоян из GSC за ее поддержку и опыт.
Дополнительный материал
Дополнительный материал к этой статье можно найти в Интернете по адресу: https://www. frontiersin.org/articles/10.3389/fmicb.2020.580719/full#supplementary-material
frontiersin.org/articles/10.3389/fmicb.2020.580719/full#supplementary-material
Дополнительный рисунок 1 | Циркулярный аннотированный Cardiosporidium cionae Апикопласт. Были извлечены два апикопласта C. cionae , число которых составляло 99.55% похожи в целом и содержат идентичную организацию генов, с SNP, локализованными в гене sufB. Апикопласты C. cionae сходны по размеру, организации и содержанию генов с апикопластами Nephromyces .
Дополнительный рисунок 2 | Сравнение геномов бактериальных эндосимбионтов Cardiosporidium cionae (AlphaC) и Nephromyces (AlphaN, Beta, Bac). Левая диаграмма Венна изображает ортологичные группы, предсказанные OrthoFinder, а правая показывает функциональное перекрытие, предсказанное с помощью KEGG.
Дополнительный рисунок 3 | Глобальная метаболическая карта генома Cardiosporidium cionae и его α-протеобактериального эндосимбионта, с путями, исключительными для C. cionae , показанными синим цветом, исключительными для α-эндосимбионта, показанными красным, и перекрывающимися метаболическими путями, показанными желтым цветом. Этот рисунок был создан с помощью iPath 3 (Darzi et al., 2018).
cionae , показанными синим цветом, исключительными для α-эндосимбионта, показанными красным, и перекрывающимися метаболическими путями, показанными желтым цветом. Этот рисунок был создан с помощью iPath 3 (Darzi et al., 2018).
Дополнительная таблица 1 | Близкородственные эталонные геномы, используемые для аннотации Cardiosporidium cionae α-эндосимбионта, загружены из GenBank, refseq.
Дополнительная таблица 2 | Аннотации ключевых функциональных генов из генома и транскриптома Cardiosporidium cionae , соответствующие показанным на рисунке 2, начиная с верхнего среднего среза и двигаясь по часовой стрелке. Аннотации основаны на функциональных прогнозах от KEGG и ссылаются на соответствующий K-число.
Каталожные номера
Абрахамсен, М.С., Темплтон, Т.Дж., Эномото, С., Абраханте, Дж.Е., Чжу, Г., Ланкто, К.А., и соавт.(2004). Полная последовательность генома апикомплексана, Cryptosporidium parvum . Наука 304, 441–445. doi: 10.1126/наука.1094786
doi: 10.1126/наука.1094786
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Аларкон, М. Э., Хара-Ф, А., Брионес, Р. К., Дубей, А. К., и Сламовиц, С. Х. (2017). Заражение грегарином ускоряет личиночное развитие кошачьей блохи Ctenocephalides Felis (Bouché). Паразитология 144, 419–425. дои: 10.1017/S0031182016002122
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Банкевич А., Нурк С., Антипов Д., Гуревич А.А., Дворкин М., Куликов А.С., и соавт. (2012). SPAdes: новый алгоритм сборки генома и его приложения для секвенирования отдельных клеток. Дж. Вычисл. биол. 19, 455–477. doi: 10.1089/cmb.2012.0021
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Капелла-Гутьеррес, С., Силья-Мартинес, Х.М., и Габальдон, Т. (2009). TrimAl: инструмент для автоматической обрезки выравнивания в крупномасштабном филогенетическом анализе. Биоинформатика 25, 1972–1973. doi: 10.1093/биоинформатика/btp348
doi: 10.1093/биоинформатика/btp348
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Чен, Х., и Бутрос, П. К. (2011). VennDiagram: пакет для создания диаграмм Венна и Эйлера с широкими возможностями настройки в R. BMC Bioinformatics 12:35. дои: 10.1186/1471-2105-12-35
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Чансио А., Сциппа С., Финетти-Сиалер М., Де Кандиа А., Аваллоне Б. и Де Винсентиис М.(2008). Переописание Cardiosporidium cionae (Van Gaver and Stephan, 1907) (Apicomplexa: Piroplasmida), плазмодиального паразита гемоцитов асцидий. евро. Дж. Протистол. 44, 181–196. doi: 10.1016/j.ejop.2007.11.005
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Конрад, П. А., Миллер, М. А., Кройдер, К., Джеймс, Э. Р., Мазет, Дж., Дабриц, Х., и соавт. (2005). Передача токсоплазмы: подсказки, полученные в результате изучения морских выдр как часовых Toxoplasma gondii , попадают в морскую среду. Междунар. Дж. Паразитол. 35, 1155–1168. doi: 10.1016/j.ijpara.2005.07.002
Междунар. Дж. Паразитол. 35, 1155–1168. doi: 10.1016/j.ijpara.2005.07.002
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Кроуфорд, М. Дж., Томсен-Зигер, Н., Рэй, М., Шахтнер, Дж., Роос, Д. С., и Сибер, Ф. (2006). Toxoplasma gondii удаляет липоевую кислоту хозяина, несмотря на ее синтез de novo в апикопласте. EMBO J. 25, 3214–3222. doi: 10.1038/sj.emboj.7601189
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Криадо-Форнелио, А., Verdú-Expósito, C., Martin-Pérez, T., Heredero-Bermejo, I., Pérez-Serrano, J., Guàrdia-Valle, L., et al. (2017). Исследование грегарин (Protozoa: Apicomplexa) у членистоногих в Испании. Паразитол. Рез. 116, 99–110. doi: 10.1007/s00436-016-5266-0
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Дайсон, Дж., Грэм, Дж., и Эвеннетт, П.Дж. (1993). Мукрон грегарина Digyalum oweni (Protozoa: Apicomplexa), паразитирующего на видах Littorina (Mollusca: Gastropoda). J. Nat. История 27, 557–564. дои: 10.1080/00222939300770311
J. Nat. История 27, 557–564. дои: 10.1080/00222939300770311
Полнотекстовая перекрестная ссылка | Академия Google
Фитцпатрик, Д. А., Криви, С. Дж., и Макинерни, Дж. О. (2006). Филогенез генома указывает на значимую филогению α-протеобактерий и поддерживает группировку митохондрий с риккетсиалами. Мол. биол. Эвол. 23, 74–85. doi: 10.1093/molbev/msj009
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Фредриксон, Дж.К., Захара Дж. М., Балквилл Д. Л., Кеннеди Д., Ли С. М., Костандаритес Х. М. и соавт. (2004). Геомикробиология аэрационных отложений, загрязненных ядерными отходами, на полигоне Хэнфорд, штат Вашингтон. Заяв. Окружающая среда. микробиол. 70, 4230–4241. doi: 10.1128/AEM.70.7.4230-4241.2004
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Френаль, К., Дубремец, Ж.-Ф., Лебрен, М., и Солдати-Фавр, Д. (2017). Скользящая подвижность обеспечивает вторжение и выход у Apicomplexa. Нац. Преподобный Микробиолог. 15, 645–660. doi: 10.1038/nrmicro.2017.86
Нац. Преподобный Микробиолог. 15, 645–660. doi: 10.1038/nrmicro.2017.86
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Гюнтер, С., Макмиллан, П.Дж., Уоллес, Л.Дж.М., и Мюллер, С. (2005). Plasmodium falciparum обладает специфическими для органелл комплексами α-кетокислоты дегидрогеназы и путями липоилирования. Биохим. соц. Транс. 33, 977–980. дои: 10.1042/BST0330977
Полнотекстовая перекрестная ссылка | Академия Google
Гюнтер, С., Уоллес, Л., Patzewitz, E.-M., McMillan, P.J., Storm, J., Wrenger, C., et al. (2007). Протеинлигаза B липоевой кислоты апикопласта не является существенной для Plasmodium falciparum . PLoS Патог. 3:e189. doi: 10.1371/journal.ppat.0030189
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Хаас, Б. Дж., Папаниколау, А., Яссур, М., Грабхер, М., Блад, П. Д., Боуден, Дж., и соавт. (2013). Реконструкция последовательности транскрипта de novo из RNA-Seq с использованием платформы trinity для создания эталонов и анализа. Нац. протокол 8, 1494–1512. doi: 10.1038/nprot.2013.084
Нац. протокол 8, 1494–1512. doi: 10.1038/nprot.2013.084
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Хофф, К. Дж., и Станке, М. (2013). WebAUGUSTUS — веб-сервис для обучения AUGUSTUS и предсказания генов у эукариот. Рез. нуклеиновых кислот. 41, W123–W128. doi: 10.1093/nar/gkt418
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Холт, К., и Янделл, М. (2011). MAKER2: конвейер аннотаций и инструмент управления базой данных геномов для проектов генома второго поколения. Биоинформатика BMC 12:491. дои: 10.1186/1471-2105-12-491
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Hyatt, D., Chen, G.-L., LoCascio, P.F., Land, M.L., Larimer, F.W., and Hauser, L.J. (2010). Prodigal: распознавание прокариотических генов и идентификация сайта инициации трансляции. Биоинформатика BMC 11:119. дои: 10.1186/1471-2105-11-119
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Джериссен, С. H., Janse, E.M., Vermeulen, A.N., и Vervelde, L. (1996). Инфекции Eimeria tenella у кур: аспекты хозяин-паразит: взаимодействие. Вет. Иммунол. Иммунопатол.
H., Janse, E.M., Vermeulen, A.N., и Vervelde, L. (1996). Инфекции Eimeria tenella у кур: аспекты хозяин-паразит: взаимодействие. Вет. Иммунол. Иммунопатол.
Полнотекстовая перекрестная ссылка | Академия Google
Джонсон, Дж., и Эльмюллер, Р. (2009). Мутуализм или паразитизм: жизнь в нестабильном континууме. Что мы можем узнать из мутуалистического взаимодействия между Piri-formospora indica и Arabidopsis thaliana? – Обзор. Endocytobiosis Cell Res. 19, 81–111.
Академия Google
Канехиса М., Фурумичи М., Танабэ М., Сато Ю. и Моришима К. (2017). KEGG: новые взгляды на геномы, пути, болезни и лекарства. Рез. нуклеиновых кислот. 45, Д353–Д361. дои: 10.1093/нар/gkw1092
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Канг, Д. Д., Фрула, Дж., Иган, Р., и Ван, З. (2015). MetaBAT, эффективный инструмент для точной реконструкции отдельных геномов из сложных микробных сообществ. PeerJ 3:e1165. doi: 10.7717/peerj.1165
PeerJ 3:e1165. doi: 10.7717/peerj.1165
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Като, К., и Стэндли, Д.М. (2013). Программное обеспечение MAFFT для множественного выравнивания последовательностей, версия 7: улучшения производительности и удобства использования. Мол. биол. Эвол. 30, 772–780. doi: 10.1093/molbev/mst010
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Козек, В. Дж., и Рао, Р. У. (2007). «Обнаружение вольбахии у членистоногих и нематод — историческая перспектива», в Wolbachia: жизнь жука в другом жуке , Vol.5, ред. А. Хоерауф и Р. У. Рао (Базель: Karger Publishers), 1–14. дои: 10.1159/000104228
Полнотекстовая перекрестная ссылка | Академия Google
Krzywinski, M., Schein, J., Birol, I., Connors, J., Gascoyne, R., Horsman, D., et al. (2009). Circos: информационная эстетика для сравнительной геномики. Рез. генома. 19, 1639–1645. doi: 10. 1101/gr.092759.109
1101/gr.092759.109
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Квонг, В.К., дель Кампо, Дж., Матур, В., Вермейдж, М.Дж. А. и Килинг, П. Дж. (2019). Широко распространенный апикомплексан, заражающий кораллы, с генами биосинтеза хлорофилла. Природа 568, 103–107. doi: 10.1038/s41586-019-1072-z
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Ламберт, К.С., Ламберт, Г., Крундуэлл, Г., и Кантарджиев, К. (1998). Накопление мочевой кислоты у одиночного асцидия Corella inflata . Дж. Экспл. Зоол. 282, 323–331. doi: 10.1002/(sici)1097-010x(19981015)282:3<323::aid-jez5>3.0.co;2-o
Полнотекстовая перекрестная ссылка | Академия Google
Леунг, Т.Л.Ф., и Поулин, Р. (2008). Паразитизм, комменсализм и мутуализм: исследование многих оттенков симбиозов . Том. 58. Франция: Vie et Milieu, 107–115.
Академия Google
Мэдден, Т. (2003). Инструмент анализа последовательности BLAST. Bethesda, MD: Национальный центр биотехнологической информации.
Bethesda, MD: Национальный центр биотехнологической информации.
Академия Google
Матур, В., дель Кампо, Дж., Колиско, М.и Килинг, П.Дж. (2018). Мировое разнообразие и распространение близких родственников апикомплексных паразитов. Окружающая среда. микробиол. 20, 2824–2833. дои: 10.1111/1462-2920.14134
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Матур, В., Колиско, М., Хеенбергер, Э., Ирвин, Н.А.Т., Леандер, Б.С., Кристмундссон, А., и соавт. (2019). Множественное независимое происхождение апикомплексаноподобных паразитов. Курс. биол. 29, 2936–2941.e5. doi: 10.1016/j.cub.2019.07.019
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Митчелл, А.Л., Эттвуд, Т.К., Бэббит, П.С., Блюм, М., Борк, П., Бридж, А., и другие. (2019). InterPro в 2019 году: улучшение охвата, классификации и доступа к аннотациям последовательностей белков. Рез. нуклеиновых кислот. 47, Д351–Д360. doi: 10.1093/nar/gky1100
doi: 10.1093/nar/gky1100
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Молл К., Юнгстрём И., Перлманн Х., Шерф А. и Вальгрен М.(2008). Методы исследования малярии . Том. 5. Центр исследований и эталонных реагентов малярии (MR4), 351.
Академия Google
Муньос-Гомес, С.А., Дурнин, К., Эме, Л., Пейт, К., Лейн, К.Е., Саффо, М.Б., и соавт. (2019а). Nephromyces представляет собой разнообразную и новую линию Apicomplexa, которая сохранила апикопласты. Геном Биол. Эвол. 11, 2727–2740. doi: 10.1093/gbe/evz155
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Муньос-Гомес, С.A., Hess S., Burger G., Lang B.F., Susko E., Slamovits C.H., et al. (2019б). Обновленная филогения Alphaproteobacteria показывает, что паразитические Rickettsiales и Holosporales имеют независимое происхождение. eLife 8:e42535. doi: 10.7554/eLife.42535
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Накаяма К. , Курокава К., Фукухара М., Ураками Х., Ямамото С., Ямадзаки К. и др. (2010). Сравнение геномов и филогенетический анализ штаммов Orientia tsutsugamushi . Рез. ДНК. 17, 281–291. doi: 10.1093/dnares/dsq018
, Курокава К., Фукухара М., Ураками Х., Ямамото С., Ямадзаки К. и др. (2010). Сравнение геномов и филогенетический анализ штаммов Orientia tsutsugamushi . Рез. ДНК. 17, 281–291. doi: 10.1093/dnares/dsq018
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Нгуен, Л.-Т., Шмидт, Х.А., фон Хазелер, А., и Мин, Б.К. (2015). IQ-TREE: быстрый и эффективный стохастический алгоритм для оценки филогений максимального правдоподобия. Мол. биол. Эвол. 32, 268–274. doi: 10.1093/molbev/msu300
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Нолфи, Дж. Р. (1970). Биосинтез мочевой кислоты в оболочниках, Molgula manhattensis , с общей схемой функции запасенных пуринов у животных. Комп. Биохим. Физиол. 35, 827–842. doi: 10.1016/0010-406x(70)-2
Полнотекстовая перекрестная ссылка | Академия Google
Пэйт, К., Хантер, Э.С., и Лейн, К.Э. (2020). Созависимость в рое видов Nephromyces зависит от гетероспецифических бактериальных эндосимбионтов. BioRxiv [Препринт] doi: 10.1101/2020.10.18.344572v1
BioRxiv [Препринт] doi: 10.1101/2020.10.18.344572v1
Полнотекстовая перекрестная ссылка | Академия Google
Пэйт, К., Сламовиц, К. Х., Саффо, М. Б., и Лейн, К.Э. (2019). Nephromyces кодирует путь метаболизма уратов и предсказал пероксисомы, демонстрируя, что это не древние потери апикомплексов. Геном Биол. Эвол. 11, 41–53. doi: 10.1093/gbe/evy251
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Паппас, Г., Руссос, Н., и Фалагас, М.Э. (2009). Моментальные снимки токсоплазмоза: глобальный статус Toxoplasma gondii , серораспространенность и последствия для беременности и врожденного токсоплазмоза. Междунар. Дж. Паразитол. 39, 1385–1394. doi: 10.1016/j.ijpara.2009.04.003
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Паркс, Д. Х., Имельфорт, М., Скеннертон, К. Т., Хугенхольц, П., и Тайсон, Г. В. (2015). CheckM: оценка качества микробных геномов, восстановленных из изолятов, отдельных клеток и метагеномов. Рез. генома. 25, 1043–1055. doi: 10.1101/гр.186072.114
Рез. генома. 25, 1043–1055. doi: 10.1101/гр.186072.114
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Ричи, М.E., Phipson, B., Wu, D., Hu, Y., Law, C.W., Shi, W., et al. (2015). Limma обеспечивает анализ дифференциальной экспрессии для секвенирования РНК и исследований микрочипов. Рез. нуклеиновых кислот. 43:e47. doi: 10.1093/nar/gkv007
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Родригес-Р., Л.М., Гунтуру, С., Харви, В.Т., Росселло-Мора, Р., Тиедже, Дж.М., Коул, Дж.Р., и соавт. (2018). Веб-сервер атласа микробных геномов (MiGA): анализ таксономического и генного разнообразия архей и бактерий на уровне всего генома. Рез. нуклеиновых кислот. 46, W282–W288. doi: 10.1093/nar/gky467
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Родригес-Р., Л. М., и Константинидис, К. Т. (2016). Коллекция enveomics: набор инструментов для специализированного анализа микробных геномов и метагеномов. PeerJ Prepr. 4:e1900v1. doi: 10.7287/peerj.preprints.1900v1
PeerJ Prepr. 4:e1900v1. doi: 10.7287/peerj.preprints.1900v1
Полнотекстовая перекрестная ссылка | Академия Google
Рюкерт, С., Уэйкман, К. С., Дженке-Кодама, Х., и Леандер, Б.С. (2015). Молекулярная систематика морских грегариновых апикомплексов из тихоокеанских оболочников с описанием пяти новых видов Lankesteria . Междунар. Дж. Сист. Эвол. микробиол. 65, 2598–2614. doi: 10.1099/ijs.0.000300
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Саффо М.Б., Маккой А.М., Рикен С. и Сламовиц С.Х. (2010). Nephromyces , полезный симбионт апикомплекса морских животных. Проц. Натл. акад.науч. США 107, 16190–16195. doi: 10.1073/pnas.1002335107
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Саффо, М.Б., и Нельсон, Р. (1983). Клетки Nephromyces : стадии развития одного жизненного цикла. Кан. Дж. Бот. 61, 3230–3239. дои: 10.1139/b83-360
Полнотекстовая перекрестная ссылка | Академия Google
Сибли, Л. Д., и Бутройд, Дж. К. (1992). Построение молекулярного кариотипа для Toxoplasma gondii . Мол. Биохим. Паразитол. 51, 291–300. дои: 10.1016/0166-6851(92)-й
Д., и Бутройд, Дж. К. (1992). Построение молекулярного кариотипа для Toxoplasma gondii . Мол. Биохим. Паразитол. 51, 291–300. дои: 10.1016/0166-6851(92)-й
Полнотекстовая перекрестная ссылка | Академия Google
Симао, Ф. А., Уотерхаус, Р. М., Иоаннидис, П., Кривенцева, Е. В., и Здобнов, Е. М. (2015). BUSCO: оценка сборки генома и полноты аннотации с помощью однокопийных ортологов. Биоинформатика 31, 3210–3212. doi: 10.1093/биоинформатика/btv351
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Солдати, Д., Дубремец, Дж. Ф., и Лебрен, М. (2001). Белки микронемы: структурные и функциональные требования для обеспечения адгезии и инвазии апикомплексанового паразита Toxoplasma gondii . Междунар. Дж. Паразитол. 31, 1293–1302. doi: 10.1016/s0020-7519(01)00257-0
Полнотекстовая перекрестная ссылка | Академия Google
Соу, С.О., Мухсен, К., Насрин, Д., Блэквелдер, В.К., Ву, Ю. , Фараг, Т.Х., и соавт. (2016). Бремя диарейной болезни Cryptosporidium среди детей в возрасте до 24 месяцев в регионах с умеренным/высоким уровнем смертности в странах Африки к югу от Сахары и Южной Азии с использованием данных глобального многоцентрового исследования кишечных инфекций (GEMS). PLoS Негл. Троп. Дис. 10:e0004729. doi: 10.1371/journal.pntd.0004729
, Фараг, Т.Х., и соавт. (2016). Бремя диарейной болезни Cryptosporidium среди детей в возрасте до 24 месяцев в регионах с умеренным/высоким уровнем смертности в странах Африки к югу от Сахары и Южной Азии с использованием данных глобального многоцентрового исследования кишечных инфекций (GEMS). PLoS Негл. Троп. Дис. 10:e0004729. doi: 10.1371/journal.pntd.0004729
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Суджа Г., Крипа В., Мохамед К., Потоди С. и Санил Н. К. (2016). Нематопсис Sp. (Apicomplexa: Porosporidae) инфекция Crassostrea madrasensis и связанная с ней гистопатология. J. Mar. Biol. доц. Индия 58, 29–33. дои: 10.6024/jmbai.2016.58.1.1890-04
Полнотекстовая перекрестная ссылка | Академия Google
Тимошенко С., Оппенгейм, Р. Д., Агрен, Р., Нильсен, Дж., Солдати-Фавр, Д., и Хациманикатис, В. (2015). Метаболические потребности и возможности Toxoplasma gondii посредством комбинированного вычислительного и экспериментального анализа. Вычисление PLoS. биол. 11:e1004261. doi: 10.1371/journal.pcbi.1004261
Вычисление PLoS. биол. 11:e1004261. doi: 10.1371/journal.pcbi.1004261
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
фон Мейенфельдт, Ф.А.Б., Архипова, К., Камбуй, Д.Д., Коутиньо, Ф.Х., и Дутиль, Б.Е. (2019). Надежная таксономическая классификация неизведанных микробных последовательностей и ячеек с CAT и BAT: дополнительная таблица 1. BioRxiv [Препринт] doi: 10.1101/530188
Полнотекстовая перекрестная ссылка | Академия Google
Вотыпка Й., Модри Д., Оборник М., Шлапета Й. и Лукеш Дж. (2016). «Apicomplexa», в Handbook of the Protists , под редакцией Дж. М. Арчибальда, А. Г. Б. Симпсона, Ч. Х. Сламовица, Л. Маргулиса, М. Мелконяна, Д. Дж. Чепмена и др. (Чам: Springer International Publishing), 1–58. дои: 10.1007/978-3-319-32669-6_20-1
Полнотекстовая перекрестная ссылка | Академия Google
Вик, Р.Р., Шульц М.Б., Зобель Дж. и Холт К.Е. (2015). Bandage: интерактивная визуализация геномных сборок de novo : рис. 1. Биоинформатика 31, 3350–3352. doi: 10.1093/биоинформатика/btv383
1. Биоинформатика 31, 3350–3352. doi: 10.1093/биоинформатика/btv383
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Ву, Ю. Х., Ансари, Х., Отто, Т. Д., Клингер, К. М., Колиско, М., Михалек, Дж., и другие. (2015). Геномы хромрид раскрывают эволюционный путь от фотосинтезирующих водорослей к облигатным внутриклеточным паразитам. eLife 4:e06974. doi: 10.7554/eLife.06974
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Юрченко Т., Шевчикова Т., Пржибыл П., Эль Каркури К., Климеш В., Амарал Р. и др. (2018). Событие переноса генов предполагает долгосрочное партнерство между эустигматофитными водорослями и новой линией эндосимбиотических бактерий. ISME J. 12, 2163–2175. doi: 10.1038/s41396-018-0177-y
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Голландцы опасаются угрозы либерализму из-за «легких наркотиков» .
Голландка курит марихуану в кофейне в Амстердаме 6 октября 2011 года. REUTERS/Toussaint Kluiters подавление рассматривается как еще одно свидетельство эрозии толерантности в стране, известной своей либеральной социальной политикой.
REUTERS/Toussaint Kluiters подавление рассматривается как еще одно свидетельство эрозии толерантности в стране, известной своей либеральной социальной политикой.
Стремление пресечь легкие наркотики исходит в основном от христианских демократов, младшего партнера в правительстве меньшинства и одной из крупных партий в условиях фрагментированного политического ландшафта.
«Очевидно, что в дебатах о морали произошел сдвиг. Все дело в культуре контроля», — сказал Дирк Корф, профессор криминологии Амстердамского университета.
Мгновенно узнаваемые по приторно-сладкому запаху жженых листьев, которые доносятся до улицы, всемирно известные «кофейни» Нидерландов почти так же распространены, как супермаркеты в больших городах, таких как Амстердам и Роттердам, и в некоторых приграничных городах.
Подобно обученным сомелье, сотрудники или «заказчики бутонов» являются экспертами по вкусу и последствиям всего, что есть в меню — белая вдова, ванильный куш или хейзеры, такие как амнезия, «известная своим экстремальным, почти параноидальным психоделическим эффектом». , с незабываемым сильным фруктовым вкусом и запахом.
, с незабываемым сильным фруктовым вкусом и запахом.
Работники стойки бойко торгуют пластиковыми пакетиками с травой, готовыми косяками и кусочками гашиша для тех, кто хочет взять с собой.
В Нидерландах допускается продажа до 5 граммов марихуаны и гашиша на человека в день в контролируемой среде кафе. Он также допускает выращивание растений марихуаны в домашних условиях в пределах пяти растений на человека, но любое выращивание большего размера является незаконным.
Высокий спрос породил секретные плантации каннабиса, которые обеспечивают так называемые запасные поставки в кофейни и являются головной болью для голландских властей, которым приходится их находить и нападать на них.
НАРКОТИКИ ТУРИСТЫ
Обычным субботним вечером кофейни в центре Амстердама забиты курильщиками. Клиентура — средний класс, голоса в основном иностранные — итальянцы, испанцы, французы, немцы, англичане.
Обеспокоенная этим наплывом туристов, употребляющих легкие наркотики, не говоря уже о том, что она считает связанной с этим преступностью, неудобствами и рисками для здоровья, Христианско-демократическая партия хочет закрыть около 700 кофеен в стране, но на данный момент согласие на введение ограничений на их деятельность.
Законопроект, который, как ожидается, будет принят парламентом к концу этого года, предусматривает, что кофейни будут работать как клубы только для членов, а это означает, что только местные жители будут иметь право регистрироваться для получения «пропусков на травку», что фактически запрещает иностранцам покупать легкие наркотики. .
В некоторых городах уже ввели более жесткие ограничения, ограничив близость кафе к школам или перенеся их на окраины. 1 октября кофейни в юго-восточном городе Маастрихт запретили вход всем иностранцам, кроме соседних немцев и бельгийцев, в качестве первого шага к введению пропусков на травку.
Эксперт по уголовным делам Корф говорит, что для запрета мало оправданий, а доказательств того, что голландская общественность поддерживает это изменение, мало.
«Серьезных опросов не проводилось, мы не знаем, изменились ли вообще мнения о кофейнях», — сказал Корф.
«До кофеен у нас была уличная торговля, они продавали на улице марихуану и обворовывали туристов. Вся проблема с наркотиками ничто по сравнению с (тем, что было) в 1980-х, 1990-х — у нас нет проблемы с героином.
Вся проблема с наркотиками ничто по сравнению с (тем, что было) в 1980-х, 1990-х — у нас нет проблемы с героином.
Институт Тримбос, изучающий зависимость и психическое здоровье, сообщил, что 5 процентов граждан Нидерландов курили травку или гашиш в прошлом году, по сравнению со средним показателем по ЕС, составляющим 7 процентов.
ГЛОБАЛЬНАЯ ПУТАНИЦА
Политики во всем мире ищут свежие идеи о том, как бороться со злоупотреблением наркотиками, открывая дебаты о политике в отношении легких наркотиков.
В июне группа высокопоставленных мировых лидеров объявила «войну с наркотиками» провалом и призвала правительства рассмотреть вопрос о декриминализации наркотиков, чтобы сократить потребление и ослабить власть организованной преступности.
Глобальная комиссия по наркополитике, в которую входят бывший президент Бразилии Фернандо Энрике Кардосо, бывший генеральный секретарь ООН Кофи Аннан, бывший председатель Федеральной резервной системы США Пол Волкер и британский миллиардер Ричард Брэнсон, заявила о многолетней стратегии запрета наркотиков и тюремное заключение пользователей во время борьбы с наркокартелями не сработало.
Он рекомендовал правительствам экспериментировать с правовым регулированием наркотиков, особенно каннабиса, сославшись на успехи в таких странах, как Нидерланды, Португалия и Швейцария, где потребление наркотиков сократилось.
Португалия, например, пошла намного дальше, чем Нидерланды, декриминализировав все наркотики, заменив тюремное заключение консультированием и лечением.
Христианские демократы не согласны и говорят, что политика Нидерландов негативно влияет на общественное здравоохранение и преступность.
«В других странах толерантности нет. Голландские кофейни привлекают много иностранных наркотуристов, особенно в приграничных районах, что доставляет много неудобств», — говорится в заявлении, опубликованном на сайте Христианско-демократической партии.
Центристская партия поставила под сомнение обоснованность разрешения кофеен, которое должно было разделить рынки легких и тяжелых наркотиков, и заявила, что люди, которые курят каннабис, часто обращаются к другим наркотикам.
В нем также утверждается, что действующее вещество каннабиса намного сильнее, чем двадцать лет назад, что ставит его в один ряд с более сильными наркотиками — отражение многолетнего выращивания новых сортов производителями.
Голландская комиссия ранее в этом году обнаружила, что гашиш и марихуана, продаваемые в Нидерландах, содержат около 18 процентов ТГК, основного психоактивного вещества, и заявила, что уровень выше 15 процентов ставит эти наркотики в один ряд с героином или кокаином.
Максим Верхаген, христианский демократ, который является заместителем премьер-министра, заявил 7 октября, что правительство запретит продажу каннабиса, концентрация ТГК в котором превышает 15 процентов.
Христианские демократы также хотят ужесточить правила в отношении так называемых плантаций каннабиса.
Помимо нелегальных поставок в кофейни, «большая часть незаконно выращиваемого в Нидерландах каннабиса экспортируется за границу. Существует разветвленная сеть, незаконно созданная в тисках организованной преступности», — говорится в заявлении партии.
Власти Нидерландов уже выделяют значительные ресурсы на поиск этих крупных плантаций.
Полиция работает с местной электроэнергетической компанией для выявления необычных моделей потребления, например круглосуточного использования в сараях и на чердаках, и использует крошечные вертолеты-ищейки, которые могут обнаруживать запах горшечных растений, доносящийся из вентиляционных шахт и дымоходов. , по сообщениям СМИ.
Городской совет Роттердама недавно раздал домохозяйствам «скретч-карты» в надежде, что заинтересованные граждане предупредят полицию, если узнают запах незаконных плантаций каннабиса по соседству.
БОРЬБА НА ДОМУ
Существует много оппозиции репрессиям. Голландские курильщики не приветствуют идею регистрации для получения пропусков на травку.
«Многие из моих клиентов — местные жители, художники, писатели, врачи, юристы, профессионалы. Они не хотят, чтобы их имя было в реестре — они не знают, кто может его увидеть или использовать. Поэтому они могут пойти к другим источникам на улице», — сказала Паула Батен, менеджер кофейни Siberie в центре Амстердама.
Поэтому они могут пойти к другим источникам на улице», — сказала Паула Батен, менеджер кофейни Siberie в центре Амстердама.
«Это правительство более христианское, более правое.Они не хотят наркотиков, но забывают о влиянии алкоголя».
Уже ходят разговоры о том, как иностранцы могут обойти новые правила, например, попросив граждан Нидерландов покупать легкие наркотики от их имени на вынос, и опасаются, что торговля легкими наркотиками выйдет на улицу.
Некоторые политики выступают против предложений. Эберхард ван дер Лаан, мэр Амстердама, говорит, что ограничение деятельности кофеен приведет к большему риску для здоровья, неудобствам и торговле наркотиками на улицах.Как мэр, он мог просто не применять правила пропуска сорняков.
«В настоящее время мэр проводит совещание с министром, чтобы убедить его, что меры, касающиеся кофеен, будут контрпродуктивными для Амстердама», — говорится в заявлении мэрии для Reuters.
Другие ссылаются на вероятные экономические последствия.
Нидерланды, как и другие европейские страны, были вынуждены ввести меры жесткой экономии и сократить расходы после кредитного кризиса, когда они вложили 40 миллиардов евро в спасение финансовых учреждений.
Налоговые поступления от кофеен оцениваются примерно в 400 миллионов евро в год. Согласно исследованиям министерства финансов и академических кругов, если Нидерланды легализуют «черные» поставки, выведя их «через борт», они смогут получать дополнительно 400–850 млн евро в год, включая экономию на стоимости правоохранительные органы.
Еще есть доход от туризма.
В Маастрихте, куда приезжает много дневных туристов, поскольку он находится так близко к границам Германии и Бельгии, исследование, проведенное по заказу ассоциации владельцев кофеен, подсчитало, что посетители городских кофеен тратят около 119 миллионов евро в год. в основном на покупки и питание вне дома.
Исследование, проведенное профессором Корфом из Университета Амстердама, показало, что туристы, которые посещали кофейни в центре Амстердама, имели такие же покупательские привычки, как и другие туристы, и с такой же вероятностью тратили 200 евро и более на гостиничный номер или раскошелились на шикарные рестораны или ночные клубы.