Кондиционеры веко: Кондиционер Beko BRH 095 (BRH 096) – официальный производитель
| Модель | BRH 095 (BRH 096) |
| Цвет | Белый |
| SAP код | 8805173200 |
| Код EAN | 8690842222290 |
| Размеры | |
|---|---|
| Упакованный внутренний блок (ШхГхВ) | 810×295×370 мм |
| Упакованный наружный блок (ШхГхВ) | 842×357×560 мм |
| Вес внутреннего блока – в упаковке | |
| Вес наружного блока – в упаковке | 28. 5
кг 5
кг |
| Количество загрузки контейнера 40’HQ (с комплектом) | 264 |
| Количество загрузки контейнера 40’GP (с комплектом) | 230 |
| 20-футовое количество загрузки контейнера (с комплектом) | 110 |
| Основные характеристики | |
| Тип | Кондиционер |
| Цвет и материал корпуса внутреннего блока | Серый Итаниуим |
| Хладагент | R410A |
| Общее количество хладагента | 600 г |
| Климат класс | T1 |
| Тип обогрева | Heat Pump |
| Подключение источника питания | Внутреннее |
| Охлаждение | 9000 BTU/ч |
| Мощность охлаждения | 2638 Вт |
| Обогрев | 10000 BTU/ч |
| Мощность обогрева | 2931 Вт |
| Потребляемая мощность при охлаждении | 3. 21 21 |
| Потребляемая мощность при обогреве | 3.61 |
| Класс энергопотребления (охлаждение) | A (92/75/EEC) |
| Класс энергопотребления (обогрев) | A (92/75/EEC) |
| Уровень шума внутреннего блока | 41/37/26 дБ |
| Уровень шума наружного блока | 56 дБ |
| Объем воздушного потока | 453/371/260 м3/ч |
| Удаление влаги | 1
L/H. r r |
| Диапазон рабочих температур окружающей среды – внешнее охлаждение | От +18 до +48 °C |
| Диапазон рабочих температур окружающей среды – внешний обогрев | От -7 до +24 °C |
| Функции и особенности | |
| Фильтры | 2 фильтра высокой плотности |
| Дополнительные фильтры | 1 Фильтр с активированным углем; 1 Фильтр с холодным катализатором |
| Дистанционное управление | LCD |
| Цифровой дисплей | VLED |
| Быстрое охлаждение | + |
| Быстрый обогрев | + |
| Функция автоматического перезапуска | + |
| Функция осушения | + |
| Автоматический контроль температуры | + |
| Ночной режим | + |
| Функция таймера | 24 ч |
| Функция размораживания | + |
| Уровни скорости вентилятора | Tурбо/H/M/L/Aвто |
| Автоматическое направление воздуха (вверх-вниз) | + |
| Функция автоматической очистки | + |
| Авто-переналадка | + |
| Функция “ZoneFollow” | + |
| Дополнительные характеристики | |
| Напряжение | 220-240 В |
| Частота тока | 50 Гц |
Компания Beko, кондиционеры Beko, сплит-системы Beko, веко
Торговая марка BEKO — часть корпорации Koç Holding Durable Goods Group.

В 1990 году Koç вышла на международный рынок и в качестве экспортного бренда бытовой техники была выбрана марка BEKO, которая вошла под крыло компании Arçelik A.Ş. вместе с маркой Arçelik. Компания выпускает весь спектр крупной и мелкой бытовой техники и входит в пятерку крупнейших производителей бытовой техники в Европе.
В общей сложности компания владеет 11 брендами, включая Grundig, Bregenz, Arctic и Blomberg и представляет свою продукцию более чем в 100 странах мира.
Техника торговой марки BEKO начала продаваться в России в 1997 году. В 1999 году было открыто представительство компании в России, а в октябре 2006 года открылся собственный завод в городе Киржач Владимирской области и запущено производство. В настоящее время бренд BEKO занимает шестое место среди производителей бытовой техники в России, он хорошо известен и уважаем. Каждые 2 секунды в мире люди покупают технику торговой марки BEKO. Компания планирует расширять производственные мощности и войти в тройку лидеров на российском рынке.
BEKO охватывает ключевые ценности: успех, понимание и преданность делу.
Особенности бренда
BEKO является внимательным, уверенным в себе, энергичным и искренним брендом.
Особое качество компании — это отражение понимания наших клиентов и их потребностей.
Атрибуты бренда
BEKO является брендом, ориентированным на потребителя, который предлагает решения, объединяющие технологичность, функциональный дизайн и эффективность.
Преимущества бренда
BEKO предоставляет интеллектуальные решения, которые помогают потребителям извлечь наибольшую пользу из повседневной жизни. В первую очередь, эти решения построены на эффективности, где BEKO помогает экономить энергию, воду и время. Кроме того, функциональный дизайн и качественные свойства продукции создают простоту в эксплуатации и позволяют более эффективно использовать пространство в вашем доме. Вот почему, когда вы покупаете продукцию BEKO, вы чувствуете, что вы сделали правильный выбор.
Награды в России:
- 2011 год.
 Премия ” Лучший Производитель бытовой техники” на Всероссийском конкурсе ” Лучшие предприятия и организации России 2011 года”
Премия ” Лучший Производитель бытовой техники” на Всероссийском конкурсе ” Лучшие предприятия и организации России 2011 года” - 2011 год. Премия «Финансово-экономический олимп» в номинации «Налогоплательщик года»
- 2011 год. Премия «За вклад в экономическое развитие России» и звание «Лидер российского бизнеса».
- 2010 год. — Премия за энергосбережение в категории «Бытовая техника». Много лет в своих разработках компания делает упор на снижение энергопотребления. Все инновации внедряются только после тщательного экотестирования. Результат — коллекция ВЕКО Green line, главная особенность которой — энергопотребление, одно из самых низких в своем сегменте.
- 2009 год. — Премия «Лучшая компания года — 2009» и звание «Социально-ответственное предприятие» » за масштабную программу, в которую входит помощь детям с различными тяжелыми заболеваниями.
- 2008 год. — Награда РБК «Компания года — 2008», как наиболее динамично развивающаяся компания на российском рынке по итогам 2008 года.
 Премия ” Лучший Производитель бытовой техники” на Всероссийском конкурсе ” Лучшие предприятия и организации России 2011 года”
Премия ” Лучший Производитель бытовой техники” на Всероссийском конкурсе ” Лучшие предприятия и организации России 2011 года”
Международные награды:
- Посудомоечная машина Beko DSN 6634 FX получила высшую награду “StiWa”
- Сушильные машины Beko DCU 8230 и Beko DPU 7340 Х были выбраны журналом “Which?” в качестве “Выгодной покупки” и “Устройства, экономящего энергию”.
- Журнал «Which?» присудил премию «Best Buy» («Лучшая покупка») 4-дверному холодильнику и 2 стиральным машинам BEKO.
- 6 премий в области технологии и дизайна «Plus X» получили холодильник, встроенный духовой шкаф, шкаф для подогрева, стиральная машина и 2 посудомоечные машины BEKO.
- Первая и единственная компания по производству «белой» бытовой техники удостоилась премии за наиболее эффективный процесс производства, т.е. «Золотого сертификата», имея восемь экологически безопасных производственных предприятий.
- Впервые в мире в отрасли «белой» бытовой техники высшую награду «TPM Special Award» («Специальная премия TPM») от JIPM (Японский институт управления производством) получил завод холодильников Eskişehir, вслед за «Excellence Award» («Премия отличия»).

мобильные напольные кондиционеры и другие разновидности, обзор модели BKP-09C и других
В современном мире человеку становится трудно представить свою жизнь без кондиционера в доме. Это и неудивительно, ведь он способен обеспечить комфорт за счет своей многофункциональности и простоты в применении. Всё чаще люди обращают внимание на продукцию компании Beko, так как она непрерывно ведет работу по улучшению качества своего продукта и расширению ассортимента, учитывая предпочтения своих покупателей.
Особенности
Кондиционеры Beko круглый год обеспечивают своим владельцам комфортный климат в доме благодаря высокому качеству продукции. Такие кондиционеры очень многофункциональны и обладают огромным количеством рабочих программ. Также продукцию данного бренда отличает превосходный дизайн. Сплит-система Beko обладает полезной функцией вентилирования помещения без возможности охлаждать или обогревать его.
Сплит-система Beko обладает полезной функцией вентилирования помещения без возможности охлаждать или обогревать его.
Техника проста в применении и отличается своей безопасностью и практичностью.
С таким кондиционером можно забыть о значительных неисправностях, устройство способно диагностировать ошибки в работе самостоятельно, выводя информацию об их наличии на дисплей.
Скорость и интенсивность работы двигателя регулируется с помощью инверторного компрессора, всё зависит от заданных параметров работы устройства. Темп замедлятся после достижения заданных условий, после чего в помещении поддерживается комфортная температура. Это помогает значительно экономить электроэнергию. Используемый тип фреона в устройстве отличается своими безопасными свойствами. Компрессоры, применяемые для работы кондиционера, способны охлаждаться и нагреваться в два раза быстрее обычной установки.
Используемый тип фреона в устройстве отличается своими безопасными свойствами. Компрессоры, применяемые для работы кондиционера, способны охлаждаться и нагреваться в два раза быстрее обычной установки.
Управлять сплит-системой можно при помощи дистанционного пульта, а также беспроводной сети Wi-Fi. Всё что для этого необходимо – скачать специальное приложение на свой смартфон.
Кондиционеры Beko обладают функцией интенсивного охлаждения Jet Cool, благодаря которой охладить помещение возможно за 30 минут. Функция Jet Heat может нагреть воздух всего лишь за один час. Чтобы создать условия для спокойного и комфортного сна, в сплит-системе используется ночной режим. Долговечность кондиционеру обеспечивает специальное антикоррозийное покрытие.
Долговечность кондиционеру обеспечивает специальное антикоррозийное покрытие.
Сплит-система Beko может эффективно очистить воздух в помещении. Этому способствует отличная система фильтрации, которую образуют следующие элементы:
- воздушный фильтр – данный механизм помогает очищать воздух от вредных примесей и пыли;
- угольный фильтр – он способен нейтрализовать неприятные запахи в помещении и защищает его от вредоносных бактерий;
- био-энзимный фильтр – его главной функцией является полная стерилизация поступающего воздуха.
Не изменяя температуру можно избавиться от повышенной влажности в помещении с помощью режима осушения. Благодаря автоматическому режиму появляется возможность настраивать движения потока воздуха. Равномерная циркуляция происходит во всех направлениях.
Еще одним полезным свойством кондиционера Beko является автоматическая работа. Если запрограммировать работу устройства на сутки, то оно четко будет придерживаться своего графика.
Если запрограммировать работу устройства на сутки, то оно четко будет придерживаться своего графика.
Достоинства и недостатки
Перед тем как приобрести любую технику, рекомендуется ознакомиться с ее преимуществами и недостатками.
Основные плюсы сплит-системы Beko.
- Бесшумная работа. Кондиционер работает очень тихо, что не позволяет отвлекаться на посторонние шумы.
- Отличная производительность. Устройство может охладить даже самое душное помещение или обогреть – холодное, при этом поддерживать фильтрацию воздуха на должном уровне, регулируя скорость потока и направление его движения.
- Быстрая установка. Монтаж кондиционера требует только базовых действий.
- Экономичность. Залог успеха кондиционеров Beko – отличные параметры и доступная цена.
- Эффективное охлаждение и обогрев. Благодаря высокой мощности сплит-система способна за короткий промежуток времени охладить или обогреть помещение.
 Благодаря высокой мощности сплит-система способна за короткий промежуток времени охладить или обогреть помещение.
Благодаря высокой мощности сплит-система способна за короткий промежуток времени охладить или обогреть помещение.Основные минусы сплит-системы Beko:
- быстрое загрязнение фильтра;
- сложность в покупке запчастей;
- в режиме максимальной мощности наблюдается повышение уровня шума;
- неравномерное охлаждение и обогрев помещения;
- функция увлажнения воздуха отсутствует.
Кондиционеры Beko имеют недостатки в работе, но их перекрывают дополнительные функции, которые позволяют увеличить срок эксплуатации техники и значительно повысить уровень ее надежности.
Виды
В ассортименте бренда фигурируют классические виды кондиционеров. Сплит-система Beko может быть мобильной и полупромышленной (ее еще называют коммерческой). Также существуют напольные кондиционеры, такую технику устанавливают в помещениях с большой площадью. Но особой популярностью пользуются кондиционеры, принадлежащие к настенному виду. Компания оснастила их инверторным и неинверторным типом управления:
Сплит-система Beko может быть мобильной и полупромышленной (ее еще называют коммерческой). Также существуют напольные кондиционеры, такую технику устанавливают в помещениях с большой площадью. Но особой популярностью пользуются кондиционеры, принадлежащие к настенному виду. Компания оснастила их инверторным и неинверторным типом управления:
- Инверторные кондиционеры Beko. Такие кондиционеры являются довольно практичными в применении, они позволяют экономить электроэнергию в доме благодаря наличию в системе блока управления и регулирующего компрессора. В случае, когда сплит-система достигла заданной температуры, отключения не происходит, устройство понижает уровень мощности своей работы, при этом температура воздуха в помещении остается неизменной. Такой тип работы свойственен и мобильным кондиционерам Beko.
- Неинверторные кондиционеры Beko. Такая система работы свойственна кондиционерам, которые только охлаждают воздух в помещении и не имеют функции нагревания. После включения устройство запускает компрессор, который, в свою очередь, стимулирует понижение температуры воздуха до заданных рабочих параметров и отключается. Модельный ряд неинверторных кондиционеров бренда Beko обладает множеством полезных свойств. Например, кондиционерам линии BXRC свойственно сохранять настройки. Это подходит тем, кто имеет проблему с частым отключением электроэнергии. Модельный ряд кондиционеров BK отличается наличием антикоррозийного покрытия корпуса. Также в своей системе они имеют антибактериальный фильтр, который позволяет обеспечивать в помещении нормальную фильтрацию воздуха и осушать его при повышенной влажности.
 Такая система работы свойственна кондиционерам, которые только охлаждают воздух в помещении и не имеют функции нагревания. После включения устройство запускает компрессор, который, в свою очередь, стимулирует понижение температуры воздуха до заданных рабочих параметров и отключается. Модельный ряд неинверторных кондиционеров бренда Beko обладает множеством полезных свойств. Например, кондиционерам линии BXRC свойственно сохранять настройки. Это подходит тем, кто имеет проблему с частым отключением электроэнергии. Модельный ряд кондиционеров BK отличается наличием антикоррозийного покрытия корпуса. Также в своей системе они имеют антибактериальный фильтр, который позволяет обеспечивать в помещении нормальную фильтрацию воздуха и осушать его при повышенной влажности.
Такая система работы свойственна кондиционерам, которые только охлаждают воздух в помещении и не имеют функции нагревания. После включения устройство запускает компрессор, который, в свою очередь, стимулирует понижение температуры воздуха до заданных рабочих параметров и отключается. Модельный ряд неинверторных кондиционеров бренда Beko обладает множеством полезных свойств. Например, кондиционерам линии BXRC свойственно сохранять настройки. Это подходит тем, кто имеет проблему с частым отключением электроэнергии. Модельный ряд кондиционеров BK отличается наличием антикоррозийного покрытия корпуса. Также в своей системе они имеют антибактериальный фильтр, который позволяет обеспечивать в помещении нормальную фильтрацию воздуха и осушать его при повышенной влажности.Модельный ряд
Модельный ряд кондиционеров торговой марки Веко отличается своим разнообразием, рассмотрим некоторые из них.
Beko BKP-09C
Данная модель кондиционера подходит тем, кто постоянно в движении. Beko BKP-09C представляет собой мобильное моноблочное переносное устройство. Устройство имеет систему самоиспарения, также предусмотрена защита от переохлаждения. При возникновении проблем, система сама способна произвести диагностику.
Благодаря наличию угольного фильтра кондиционер очищает воздух от пыли. Зимой такое устройство можно использовать в качестве вентилятора, так как оно не имеет функции нагревания воздуха. Сплит-система также имеет возможность осушать помещение, что позволяет избавиться от избыточного уровня влаги. В жаркую погоду устройство способно охладить небольшую комнату все за 10 минут. Минусом кондиционера этой модели считается высокий уровень шума – 55дБ.
Beko BKK-12C
Кондиционер этой модели во многом похож на предыдущую модель, он также относится к мобильному переносному устройству. Основным режимом является охлаждение. Дополнительный режим – вентиляция, который работает без системы охлаждения или обогрева.
Управляется устройство пультом дистанционного управления. Благодаря наличию таймера можно установить время работы сплит-системы. Также есть режим осушения. Обладает функцией запоминания настроек. Уровень шума может достигать 58 дБ, что исключает возможность работы ночью из-за большого шумового фона.
Beko BCDH 070/BCDH 071
Данные кондиционеры отлично подойдут для установки в помещениях, площадь которых не превышает 20 м2. Преимущество такой системы состоит в том, что она может как охлаждать помещение, так и обогревать его. Управлять устройством можно с помощью дистанционного пульта, который работает в диапазоне нескольких метров. Уровень шума достигает 34 дБ, что является нормальным показателем.
Кондиционер может работать в турборежиме – это позволяет устройству функционировать эффективнее. Ночной режим позволит сохранить благоприятный микроклимат даже во время сна. Фильтрационная система настроена на автоматическое самоочищение внутренних элементов, а также дезодорацию. Вручную можно задавать время работы кондиционера при помощи таймера. По истечению заданного времени устройство прекращает свою работу.
Советы по выбору
К выбору кондиционера следует подходить серьезно, так как это задача сложная и требует для каждого свой индивидуальный подход. Приобретая сплит-систему, необходимо следовать определенным правилам:
- определить оптимальную мощность кондиционера путем оценки всех параметров помещения;
- выяснить, какой тип сплит-системы подходит лучше всего, узнать мнение специалистов;
- выбрать место для установки устройства;
- получить консультацию от специалистов, если это представляется возможным.

Важным правилом при покупке любой бытовой техники, которым стоит руководствоваться, является то, что если устройство исправно будет работать первый месяц, то при правильном пользовании оно прослужит очень долго, как минимум весь гарантийный срок эксплуатации.
Стоит заметить, что между гарантийным сроком и сроком эксплуатации изделия существует большая разница: если первый может быть установлен на период 1-2 года, то второй может достигать 7 лет.
Комфортная температура для каждого является индивидуальной, рекомендуемый оптимальный показатель для всей семьи может колебаться от 21 до 24 градусов. Обычно достаточно автоматизировать режим работы сплит-системы в специальном режиме.
Правила эксплуатации
Если следовать простым рекомендациям по эксплуатации, то можно значительно продлить жизнь сплит-системе, превысив ее гарантийный срок:
- при выборе кондиционера учитывайте площадь помещения, чтобы мощность устройства соответствовала его размерам;
- чтобы не допустить сильного засорения фильтрационной системы, необходимо регулярно ее очищать;
- категорически не рекомендуется перегружать кондиционер, датчик температуры не должен превышать допустимого производителем диапазона;
- включить кондиционер с открытыми окнами будет большой ошибкой – помещение не охладиться, а ресурсы устройства будут растрачены;
- пользоваться сплит-системой «зима-лето» в режиме обогрева не рекомендуется, в случае если на улице температура ниже нуля;
- ежегодно следует производить техническое обслуживание кондиционера, а также проверять состояние охлаждающей жидкости.

Каждая модель кондиционера требует индивидуального подхода, поэтому следует также ознакомиться с руководством по эксплуатации от производителя. Самое главное – использовать кондиционер с умом, и тогда он станет незаменимым помощником в жизни и будет спасать от жары долгое время.
Обзор кондиционера Beko ждет вас в видео ниже.
Кондиционер Beko не охлаждает
Нижеперечисленные причины, возникающие в случае когда кондиционер не охлаждает, указаны от наиболее вероятной до наименее вероятной. Проверьте или протестируйте каждую причину, начиная с наиболее вероятных причин.
Проверьте или протестируйте каждую причину, начиная с наиболее вероятных причин.Причина 1 Воздушный фильтр
Если воздушный фильтр засорен, то воздух не может нормально протекать через кондиционер. Это значительно снижает охлаждающую способность кондиционера. Кроме того, если воздушный фильтр забит, то замерзнут испарительные катушки. Чтобы удалить мусор из воздушного фильтра, попробуйте его очистить. Если воздушный фильтр невозможно очистить, то замените его.
Причина 2 Катушки конденсатора загрязнены
Катушки конденсатора рассеивают тепло, когда хладагент проходит через них. Если конденсаторные катушки загрязнены, то они не смогут эффективно рассеивать тепло. По мере того, как мусор накапливается на катушках, кондиционер становится менее эффективным, в результате чего кондиционер будет работать более интенсивно для выработки охлаждения. Если катушки сильно загрязнены, то кондиционер не сможет поддерживать надлежащую температуру, и компрессор будет работать непрерывно, чтобы охладить помещение. Проверьте конденсаторные катушки, чтобы определить, загрязнены ли они. Если конденсаторные катушки загрязнены, то очистите их.
Проверьте конденсаторные катушки, чтобы определить, загрязнены ли они. Если конденсаторные катушки загрязнены, то очистите их.
Причина 3 Компрессор
Возможно, неисправен компрессор. Однако это обычно не так. Перед заменой компрессора обязательно проверяйте компоненты чаще выходящие из строя, в частности, предохранитель от перегрузки и конденсатор компрессора. Если компрессор неисправен, то его следует заменить только с привлечением лицензированного мастера.
Причина 4 Термостат
Термостат контролирует температуру воздуха. Когда температура воздуха поднимается выше заданного значения, то термостат активирует переключатель, чтобы включить вентилятор и компрессор. Если термостат неисправен, то он не сможет нормально охлаждаться. Чтобы определить, неисправен ли термостат, установите кондиционер на охлаждение, а затем используйте мультиметр, чтобы проверить термостат на непрерывность. Если непрерывность термостата нарушена, когда кондиционер включен на охлаждение, то замените термостат. (Примечание. Некоторые термостаты могут также управлять отопителем и, как результат, иметь три клеммы. Для получения дополнительной информации смотрите схему электроразводки кондиционера).
(Примечание. Некоторые термостаты могут также управлять отопителем и, как результат, иметь три клеммы. Для получения дополнительной информации смотрите схему электроразводки кондиционера).
Причина 5 Термистор
Термистор – это датчик, подключенный к плате управления, определяющий температуру воздуха. Значение сопротивления термистора изменяется вместе с температурой воздуха. Если температура воздуха повышается, то значение сопротивления будет уменьшаться. Чтобы определить, неисправен ли термистор, используйте мультиметр, чтобы проверить его на непрерывность. Если непрерывность термистора нарушена или если сопротивление термистора не изменяется при изменении температуры, то замените термистор. (Чтобы найти конкретные значения температуры и сопротивления, попробуйте узнать это из руководства для пользователя.)
Причина 6 Панель управления
Контрольная панель регулирует напряжение на двигателе компрессора и двигателе вентилятора. Если плата управления выходит из строя, то другие компоненты не будут работать должным образом или вообще не будут работать..jpg) Контрольные панели часто диагностируются ошибочно; убедитесь, что Вы проверили детали которые ломаются чаще, перед заменой платы управления. Если же Вы уверены, что все остальные компоненты работают правильно, то замените плату управления.
Контрольные панели часто диагностируются ошибочно; убедитесь, что Вы проверили детали которые ломаются чаще, перед заменой платы управления. Если же Вы уверены, что все остальные компоненты работают правильно, то замените плату управления.
Причина 7 Термостат контроля температуры
Термостат контроля температуры – это датчик, контролирующий температуру воздуха и активирующий переключатель, чтобы обеспечить питание вентилятора и компрессора, когда температура воздуха опускается ниже заданного значения. Если термостат не работает, то кондиционер будет охлаждать непрерывно или вообще не будет охлаждать. Чтобы определить, неисправен ли терморегулятор температуры, используйте мультиметр, чтобы проверить его на непрерывность. Если непрерывность термостата нарушена, то замените его. (Примечание. Некоторые термостаты могут также управлять отопителем и, как результат, иметь три клеммы. Для получения дополнительной информации смотрите схему электропроводки кондиционера).
Причина 8 Панель контроля температуры
Если панель контроля температуры неисправна, то двигатель вентилятора и компрессор не получат ток. Контрольные панели часто ошибочно диагностируются; так что убедитесь, что Вы проверили все другие детали кондиционера перед заменой платы контроля температуры. Если же Вы уверены, что все остальные компоненты работают правильно, то замените контрольную панель.
Причина: 9 Конденсатор
Конденсатор питания подключается к компрессору с помощью электрических проводов. Если конденсатор перегорает, то компрессор не будет работать. Чтобы определить, сгорел ли конденсатор, протестируйте его с помощью мультиметра. Если непрерывность конденсатор нарушена, то замените его.
Другие неполадки с кондиционером Beko
Кондиционер не охлаждает
Вентилятор кондиционера не работает
Компрессор кондиционера не работает
Кондиционер не выключается
Кондиционер громко работает или сильно шумит
Из внутреннего блока кондиционера течет вода
Пульт дистанционного управления кондиционера не работает
Замерзает теплообменник кондиционера
Коды ошибок кондиционер Beko
Видео советы мастеров по ремонту кондиционера Beko
Кондиционер веко, инструкция по применению.
 Основные характеристики кондиционеров Веко
Основные характеристики кондиционеров ВекоКондиционер веко, инструкция по применению. Для более общего ознакомления приведем вам общую инструкцию, подходящую для большинства моделей кондиционера ВЕКО. Характеристики, приведенные в этой статье, подходят практически ко всем моделям кондиционеров ВЕКО , что делает этот материал очень полезным и удобным для пользователей.
Практически все кондиционеры ВЕКО относятся к классу энергопотребления А.Коэффициент их ффективности (Energy Efficiency Rating) — это такое соотношение между потребляемой мощностью кондиционера и теплопроизводительностью. Соответствуя нормам, принятым в Европейском союзе, все кондиционеры, имеющие коэффициент энергоэффективности больше трех Вт (это равно 10.92 тепловых единиц Британии /шестьдесят минут) на один Вт используемой мощности относят к классу энергоэффективности (A). Для тех кондиционеров, которые работают в режиме обогревания, такая норма может составить три с половиной Вт (это равно 12,28 тепловых единиц/час). Кондиционеры марки Beko класса А могут потреблять меньше электроэнергии, чем кондиционеры, которые работают в подобных условиях с низкой энергоэффективностью, при соблюдении точно такой производительности.
Кондиционеры марки Beko класса А могут потреблять меньше электроэнергии, чем кондиционеры, которые работают в подобных условиях с низкой энергоэффективностью, при соблюдении точно такой производительности.
Инвертор (Invertech)
Инверторные технологии кондиционеров Beko дают возможность регулировать направление и скорость, с которой вращается двигатель компрессора системы, в зависимости от условий охлаждения или обогрева помещения. Когда достигается необходимая температура в комнате, скорость вращения двигателя компрессора сожжет снизиться, и кондиционер перейдет в режим поддержания наиболее оптимальной температуры.
Инверторные кондиционеры являются экологически безопасным оборудованием, так как в них использован газ – фреон — R410A
В простых кондиционерах ВЕКО для того, чтоб достигать необходимой температуры в помещении компрессор может регулярно включаться и выключаться. Во время таких запусков и во время остановок может расходоваться большое количество электрической энергии. В кондиционерах с инверторным управлением для того, чтобы снизить расход энергии, вместо остановок и запусков компрессора происходит регулировка скорости, с которой вращается двигатель (понижение скорости или увеличение) в соответствии с необходимыми условиями. Это дает возможность сокращать энергопотребление в сравнении с работой стандартных кондиционеров примерно на сорок четыре процента.
В кондиционерах с инверторным управлением для того, чтобы снизить расход энергии, вместо остановок и запусков компрессора происходит регулировка скорости, с которой вращается двигатель (понижение скорости или увеличение) в соответствии с необходимыми условиями. Это дает возможность сокращать энергопотребление в сравнении с работой стандартных кондиционеров примерно на сорок четыре процента.
Для того, чтобы повысить эффективность прогрева помещения кондиционерами ВЕКО нужно:
Системы кондиционирования с инверторным управлением позволяют обеспечивать стойкую эффективность обогревания даже при соблюдении температуры внешнего ниже, чем пятнадцать градусов ниже нуля.
Благодаря более широкому диапазону рабочих температур кондиционер ВЕКО довольно эффективно работает в режиме обогревания даже при самых низких температурах.
Все кондиционеры ВЕКО имеют более высокий уровень комфорта, тихо работают
Самый идеальный комфорт обеспечивается низким уровнем шума
Кондиционеры ВЕКО с инверторным управлением позволяют быстро нагревать помещение до необходимой температуры и держать ее на определенном уровне. Так, наиболее комфортные условия работу обеспечиваются для кондиционера бесперебойно.
Так, наиболее комфортные условия работу обеспечиваются для кондиционера бесперебойно.
Такие кондиционеры работают в очень широком температурном диапазоне. В первую очередь компрессор может более интенсивно работать, для достижения необходимой температуры в помещении. После этого выполняют регулировку скорости вращения компрессора для более стабильной работы климатического оборудования.
Кроме этого, все инверторные кондиционеры Beko могут работать более тихо, чем обычные кондиционеры — шум во внутреннем блоке при этом составит примерно двадцать шесть дБА.
Наличие интеллектуальной системы управления
Кондиционер ВЕКО при помощи этой функции может отслеживать температуру окружающего воздуха и переключатся автоматически между разными режимами функционирования (охлаждение/обогрев) в зависимости от выбранного направления. Это помогает вам поддержать температуру в комнате на нужном уровне.
Функция Jet Cool™
Jet Cool™ может охлаждать помещение в течении е тридцати минут
Данная функция предназначается для более быстрого и качественного охлаждения воздуха в закрытом помещении. Для того, чтобы включить данный режим будет достаточно включить его на пульте управления. После того, как вы включите функцию Jet Cool™, воздух в помещении за тридцать минут будет охлаждении до температуры минус восемнадцать градусов. Во время охлаждения, дополнительной необходимости установления времени работы не потребуется.
Для того, чтобы включить данный режим будет достаточно включить его на пульте управления. После того, как вы включите функцию Jet Cool™, воздух в помещении за тридцать минут будет охлаждении до температуры минус восемнадцать градусов. Во время охлаждения, дополнительной необходимости установления времени работы не потребуется.
Режим работы кондиционеров ВЕКО — Jet Heat™
В процессе использовании я данной функции кондиционер ВЕКО на протяжении шестидесяти минут может нагреть воздух в комнате до температуры тридцать градусов. В таком режиме вентилятор кондиционера начинает работать на самой большой скорости.
Важно! температура воздуха, который поступает из кондиционера, бывает разной в зависимости от выбранной температуры в комнате, температуры внешнего воздуха и от производительности самого кондиционера.
Режим работы ночью:
Ночью, температура человеческого тела понижается. Потому, через тридцать минут после того, как кондиционер переходи т в режим работы в ночное время кондиционер автоматически повышает температуру воздуха на один градус, а, когда пройдет еще шестьдесят минут повышает температуру еще на два градуса. Такой порядок действий позволяет создавать самые комфортные и благоприятные условия для работы кондиционера в процессе и помогает избегать чувство холода.
Такой порядок действий позволяет создавать самые комфортные и благоприятные условия для работы кондиционера в процессе и помогает избегать чувство холода.
Специальное покрытие с названием Gold Fin™
“Gold Fin™” — это такое покрытие, которое защищает кондиционер от коррозии
Поверхность конденсатора из алюминия во внешнем блоке покрыт «Gold Fin™». Это покрытие позволяет продлевать срок работы кондиционера и защищать конденсатор внешнего блока от коррозии металла.
Режим работы «Хаос» позволяет равномерно распределять поток воздуха.
В режиме работы «Хаос» вентилятор может автоматически переключаться с большой скорости на маленькую и наоборот
Режим работы кондиционера ВЕКО — безопасное осушение
В таком особом режиме осушения вы можете безопасно высушивать воздух, не сильно снижая температуру воздуха в помещении. Так , когда не очень жарко, но вам мешает повышенная влажность, есть возможность осушить воздух в комнате, при этом не сильно охлаждая воздух.
Наличие биоактивного угольного фильтра
При помощи биоактивного угольного фильтра воздух может очищаться от разных загрязнений.
Антибактериальный воздушный фильтр, который можно мыть
Такой фильтр может задерживать разные частицы, которые летают в воздухе, например, пыль или пыльца от цветов.
Наличие дополнительного режима вентиляции
В таком режиме может работать только вентилятор мобильного блока кондиционера. Ни режим обогрева, ни режим охлаждения не проводится в этот момент. В такой способ можно обеспечить правильную циркуляцию воздуха в комнате.
Наличие энергосберегающего режима
При работе в таком режиме температура, а также скорость вентилятора может устанавливаться в автоматическом режиме, в зависимости от типовых условий, более приятных для каждого человека, что позволяет обеспечивать экономное расходование электроэнергии. В таком режиме работы кондиционер во время понижении я температуры в комнате может автоматически переходить в работу в режиме обогревания, а во время повышения температуры — он переходит в режим работы на холод.
Важно! При работе в таком энергосберегающем режиме переключение между разными режимами работы кондиционера не производится.
В кондиционерах есть возможность осуществлять автоматический контроль за температурой.
В процессе работы кондиционер может постоянно измерять температуру в комнате с целью установления и поддержания температуры на заданном уровне.
Автоматическое направление воздушного потока (вверх/вниз)
Эта функция обеспечивает равномерное распределение воздуха, направляя воздушный поток вниз и наверх. Если возникнет такая необходимость вы можете вручную направлять поток воздуха в правую сторону или налево.
Можно автоматически направлять воздушный поток (есть четыре направления)
Данная функция позволяет обеспечивать более равномерное и эффективное распределение воздуха, придавая нужное направление воздуху. В тех местах, в которых ее используют может усиливаться ощущение комфортности. Помимо этого, пользователю позволяется регулировать распределения воздушных потоков при помощи пульта дистанционного управления.
Все действия, которые вы хотите провести с кондиционерами Beko, можно выполнить при помощи пульта дистанционного управления. С пультом дистанционного управления вы можете легко устанавливать необходимый режим работы, включать или выключать дополнительные функции или отключать всю систему очищения воздуха Neo Plasma™.
А еще у вас есть возможность программировать включение и выключение кондиционера в разное время на последующие двадцать четыре часа. Их отличие от стандартных кондиционеров заключается в том, что применяют два разных таймера: один таймер необходим для включения, а второй – для того, чтобы выключить кондиционер. Так, вы можете программировать отключение кондиционера, в то время, когда его работа не нужна, и таким образом экономить электрическую энергию, или его включение перед возвращением домой, чтобы создать к моменту прихода комфортную обстановку.
Наличие цифрового индикатора
На мобильном блоке имеется специальный цифровой индикатор, на котором отображаются параметры, с которыми работает кондиционер: а также указывается режим, температура работы или выбранная программа.
Почти все кондиционеры ВЕКО обладают сниженным уровнем шума.
Кондиционер Beko очень тихо работает благодаря новейшей конструкции вентилятора мобильного блока и инновационной схеме, по которой распределяется поток воздуха.
Помогла статья?
| Номер на схеме: 1 | Артикул:9192434366 | Наличие:Под заказ | Цена:670 руб | Купить | ||
| Номер на схеме: 4,5 | Артикул:9192434693 | Наличие:Под заказ | Цена:2 648 руб | Купить | ||
Номер на схеме: 5 | Артикул:9192436280 | Наличие:Под заказ | Цена:1 390 руб | Купить | ||
| Номер на схеме: 6 | Артикул:9192434191 | Наличие:Под заказ | Цена:9 981 руб | Купить | ||
| Номер на схеме: 7 | Артикул:9192434818 | Наличие:Под заказ | Цена:1 889 руб | Купить | ||
| Номер на схеме: 11 | Артикул:9192433687 | Наличие:Под заказ | Цена:2 098 руб | Купить | ||
| Номер на схеме: 12 | Артикул:9192434365 | Наличие:Под заказ | Цена:4 207 руб | Купить | ||
| Номер на схеме: 13 | Артикул:9192433721 | Наличие:Под заказ | Цена:770 руб | Купить | ||
| Номер на схеме: 13-15 | Артикул:9192434820 | Наличие:Под заказ | Цена:8 691 руб | Купить | ||
| Номер на схеме: 15,16 | Артикул:9192434692 | Наличие:Под заказ | Цена:12 587 руб | Купить | ||
| Номер на схеме: 18 | Артикул:9192434312 | Наличие:Под заказ | Цена:4 207 руб | Купить | ||
| Номер на схеме: 20 | Артикул:9192434691 | Наличие:Под заказ | Цена:539 руб | Купить | ||
| Номер на схеме: 21 | Артикул:9192434168 | Наличие:Под заказ | Цена:3 856 руб | Купить | ||
| Номер на схеме: 22 | Артикул:9192434162 | Наличие:Под заказ | Цена:12 587 руб | Купить | ||
| Номер на схеме: 23 | Артикул:9192434821 | Наличие:Под заказ | Цена:4 955 руб | Купить | ||
| Номер на схеме: 24 | Артикул:9192434822 | Наличие:Под заказ | Цена:4 207 руб | Купить | ||
| Номер на схеме: 25 | Артикул:9192433725 | Наличие:Под заказ | Цена:12 587 руб | Купить | ||
| Номер на схеме: 26,28 | Артикул:9192434186 | Наличие:Под заказ | Цена:15 005 руб | Купить | ||
| Номер на схеме: 28 | Артикул:9192433749 | Наличие:Под заказ | Цена:959 руб | Купить | ||
| Номер на схеме: 29 | Артикул:9192433748 | Наличие:Под заказ | Цена:959 руб | Купить | ||
| Номер на схеме: 99 | Артикул:9186308014 | Недоступно к заказу | ||||
| Номер на схеме: 99 | Артикул:9192434824 | Наличие:Под заказ | Цена:9 382 руб | Купить | ||
| Номер на схеме: 99 | Артикул:9192434690 | Наличие:Под заказ | Цена:340 руб | Купить | ||
| Номер на схеме: 99 | Артикул:9192434694 | Наличие:Под заказ | Цена:7 643 руб | Купить | ||
| Номер на схеме: 99 | Артикул:9192434823 | Наличие:Под заказ | Цена:1 889 руб | Купить | ||
| Номер на схеме: 99 | Артикул:9192434531 | Наличие:Под заказ | Цена:5 166 руб | Купить | ||
| Номер на схеме: 99 | Артикул:9192434696 | Наличие:Под заказ | Цена:670 руб | Купить | ||
| Номер на схеме: 99 | Артикул:9192434369 | Наличие:Под заказ | Цена:340 руб | Купить | ||
| Номер на схеме: 99 | Артикул:9192434825 | Наличие:Под заказ | Цена:9 382 руб | Купить | ||
Принадлежности для уборки | kaup24.
 ee
eeПринадлежности для уборки
Каждому из нас хочется жить и работать в уютной и чистой среде, однако для того, чтобы всегда было приятно возвращаться домой, приходится самостоятельно прилагать усилия. В данном случае помогают чистящие средства и аксессуары, позволяющие эффективно выполнять все работы по дому. Когда настанет время чистить оконные стекла, Вам помогут следующие чистящие средства: швабры и щетки для мытья с длинной телескопической ручкой, позволяющей легко доставать даже до труднодоступных мест. А если Вам потребуется помыть напольное покрытие, то помогут щетки, швабры для мытья полов или щетки для влажной чистки полов. Не менее важной является и ванная комната – в таком случае подойдут щетки для мытья ванны или швабры с ведром.
Часто используется и березовая швабра, также своей функциональностью восхищают швабры с механизмом отжима, позволяющие еще эффективнее выполнять все работы по уборке. Ни у кого не возникает сомнений по поводу того, что швабры, щетки для пола, щетки для дома – это обязательный атрибут, поэтому важно позаботиться о приобретении хотя бы нескольких из вышеперечисленных товаров.
Ни у кого не возникает сомнений по поводу того, что швабры, щетки для пола, щетки для дома – это обязательный атрибут, поэтому важно позаботиться о приобретении хотя бы нескольких из вышеперечисленных товаров.
Тех, кого интересуют аксессуары для чистящих средств, резиновые, ленточные, проволочные щетки или швабры для мытья по более низким ценам, приглашаем посетить электронный магазин kaup24.ee и поближе ознакомиться с предлагаемыми в нашем ассортименте чистящими средствами. Здесь очень привлекательные для покупателей цены на березовые швабры, но не меньше порадуют и другие предложения и удобный способ приобретения этих средств.
Когда нужна швабра для плитки, улицы или других работ, то хочется приобрести ее как можно скорее и максимально удобным способом. Теперь это сделать просто, так как швабру можно заказать через интернет всего за несколько минут, после чего мы позаботимся о том, чтобы выбранные чистящие средства в скором времени были доставлены по указанному Вами адресу. Поэтому Вам остается лишь выбрать наиболее привлекательная для Вас изделия. В ассортименте Вы найдете швабры Easy Mop, щетки York, Fiskars, Vileda, JIF, Liebherr, Leifheit, Merrilla, Vikan и щетки других производителей, чистящие средства для мытья полов и других работ по уборке. Обратите внимание на то, что в нашем электронном магазине часто проводятся акции на щетки. Во время данных акций эти и другие изделия можно приобрести по гораздо меньшим ценам.
Поэтому Вам остается лишь выбрать наиболее привлекательная для Вас изделия. В ассортименте Вы найдете швабры Easy Mop, щетки York, Fiskars, Vileda, JIF, Liebherr, Leifheit, Merrilla, Vikan и щетки других производителей, чистящие средства для мытья полов и других работ по уборке. Обратите внимание на то, что в нашем электронном магазине часто проводятся акции на щетки. Во время данных акций эти и другие изделия можно приобрести по гораздо меньшим ценам.
Добавляя, сколько нужно и когда
Abstract
Концептуальные и практические преимущества кондиционирования павловых век облегчают анализ вычислений и обучения мозжечка. Тем не менее, процедуры кондиционирования век во многом нереальны. Сигнал ошибки для оливоцеребеллярной системы не уменьшается, поскольку обучение адаптирует амплитуду или усиление ответа. Это по своей сути ограничивает полезность кондиционирования век для исследований, изучающих, как механизмы обучения мозжечка приобретают и сохраняют амплитуду адаптивного ответа. Мы сообщаем о разработке и описании процедуры обучения, в которой амплитуда условной реакции ставится под экспериментальный контроль с непредвиденными обстоятельствами, которые более близки к естественным условиям. В этой процедуре доставка безусловного стимула (US) зависит от амплитуды условного ответа: US доставляется для ответов, которые не достигают заданной целевой амплитуды, и опускается для ответов, которые соответствуют или превышают цель. Мы обнаружили, что кролики, обученные либо тональной стимуляции, либо стимуляции мшистыми волокнами в качестве условного стимула, усваивают ответы, которые достигают целевых амплитуд в диапазоне от 2 до 5 мм.Инактивация межположительного ядра с помощью инфузий мусцимола устраняет эти условные реакции, указывая на то, что участие мозжечка в кондиционировании век не связано явно с использованием процедур Павлова. Вместе с предыдущими исследованиями эти данные предполагают, что амплитуда ответа изучается и кодируется в мозжечке во время кондиционирования век.
Мы сообщаем о разработке и описании процедуры обучения, в которой амплитуда условной реакции ставится под экспериментальный контроль с непредвиденными обстоятельствами, которые более близки к естественным условиям. В этой процедуре доставка безусловного стимула (US) зависит от амплитуды условного ответа: US доставляется для ответов, которые не достигают заданной целевой амплитуды, и опускается для ответов, которые соответствуют или превышают цель. Мы обнаружили, что кролики, обученные либо тональной стимуляции, либо стимуляции мшистыми волокнами в качестве условного стимула, усваивают ответы, которые достигают целевых амплитуд в диапазоне от 2 до 5 мм.Инактивация межположительного ядра с помощью инфузий мусцимола устраняет эти условные реакции, указывая на то, что участие мозжечка в кондиционировании век не связано явно с использованием процедур Павлова. Вместе с предыдущими исследованиями эти данные предполагают, что амплитуда ответа изучается и кодируется в мозжечке во время кондиционирования век. Таким образом, эти результаты обеспечивают основу для систематических и контролируемых исследований мозжечковых механизмов, которые обучаются и кодируют правильную амплитуду адаптивных движений.
Таким образом, эти результаты обеспечивают основу для систематических и контролируемых исследований мозжечковых механизмов, которые обучаются и кодируют правильную амплитуду адаптивных движений.
Введение
Павловское кондиционирование века облегчило анализ обучения и вычислений мозжечка (McCormick and Thompson, 1984; Perrett et al., 1993; Mauk and Donegan, 1997; Medina and Mauk, 1999; Hansel et al., 2001; Medina et al., 2001, 2002), в основном из-за тесного соответствия между тренировочными стимулами и входными сигналами мозжечка, а также между выходной мощностью мозжечка и поведенческими реакциями (McCormick and Thompson, 1984; Mauk et al., 1986; Steinmetz et al., 1986; Маук и Донеган, 1997; Hesslow et al., 1999). Идея Павлова (1927) о том, что условное кондиционирование – это «рефлексивное обучение», при котором условный стимул (КС) приобретает способность активировать рефлекторный путь, запускаемый подкрепляющим стимулом, является неправильным представлением, обычно применяемым к кондиционированию век. В самом деле, неправильное название «кондиционирование моргания», по-видимому, происходит из этого заблуждения. Данные, противоречащие этой идее, включают известные схемы кондиционирования век, которые ясно показывают, что путь CS не связан с рефлекторным путем через обучение (Mauk and Donegan, 1997; Thompson et al., 1998), исходя из свидетельств того, что условные реакции век не являются морганиями (Schade Powers et al., 2010), и из простого поведенческого наблюдения, что во время приобретения амплитуда условной реакции не является однозначной. С тренировкой амплитуда условной реакции монотонно растет от едва заметного до максимального закрытия век.
В самом деле, неправильное название «кондиционирование моргания», по-видимому, происходит из этого заблуждения. Данные, противоречащие этой идее, включают известные схемы кондиционирования век, которые ясно показывают, что путь CS не связан с рефлекторным путем через обучение (Mauk and Donegan, 1997; Thompson et al., 1998), исходя из свидетельств того, что условные реакции век не являются морганиями (Schade Powers et al., 2010), и из простого поведенческого наблюдения, что во время приобретения амплитуда условной реакции не является однозначной. С тренировкой амплитуда условной реакции монотонно растет от едва заметного до максимального закрытия век.
Тем не менее, стандартные тренировочные процедуры кондиционирования век не позволяют экспериментально контролировать амплитуду ответа. Для более естественных реакций, таких как адаптация вестибулоокулярного рефлекса (VOR), ввод ошибки в оливоцеребеллярную систему (т.е.(например, на вход карабкающихся волокон в мозжечок) уменьшается по мере того, как обучение адаптирует усиление или амплитуду ответа (Gilbert and Thach, 1977; Raymond and Lisberger, 1997, 1998). Поскольку вероятность сигнала ошибки в оливоцеребеллярной системе [через предъявление безусловного стимула (УЗ)] не уменьшается по мере роста амплитуды условного ответа, процедуры Павлова по своей природе способствуют условным ответам (КО) с максимальной амплитудой (5–6 мм для кроликов). ). Даже при воздушной затяжке, такой как УЗИ, CR (расширение мигательной перепонки) не может полностью устранить активацию УЗ-лазейных волокон, потому что внешние веки обычно ограничены, а сама мигательная перепонка весьма чувствительна.По этим причинам кондиционирование век не было полезным для анализа мозжечковых механизмов, участвующих в обучении и кодировании амплитуды ответа (то есть усиления ответа).
Поскольку вероятность сигнала ошибки в оливоцеребеллярной системе [через предъявление безусловного стимула (УЗ)] не уменьшается по мере роста амплитуды условного ответа, процедуры Павлова по своей природе способствуют условным ответам (КО) с максимальной амплитудой (5–6 мм для кроликов). ). Даже при воздушной затяжке, такой как УЗИ, CR (расширение мигательной перепонки) не может полностью устранить активацию УЗ-лазейных волокон, потому что внешние веки обычно ограничены, а сама мигательная перепонка весьма чувствительна.По этим причинам кондиционирование век не было полезным для анализа мозжечковых механизмов, участвующих в обучении и кодировании амплитуды ответа (то есть усиления ответа).
Мы сообщаем о разработке и описании процедуры тренировки век, которая более точно имитирует естественные требования к обучению мозжечка и ставит амплитуду ответа под экспериментальный контроль. В этой процедуре УЗИ зависит от амплитуды условного ответа, так же как ошибочные воздействия на мозжечок зависят от усиления ответа. Когда амплитуда ответа соответствует заранее заданной целевой амплитуде или превышает ее, УЗИ не выполняется (см.). Мы демонстрируем, что амплитуда ответа приближается к целевым амплитудам в диапазоне от 2 до 5 мм, со значительными групповыми различиями при анализе после 10 тренировок. Участие мозжечка в этой обучающей задаче подтверждается двумя наблюдениями: (1) амплитуда ответа может быть определена, когда стимуляция мшистых волокон заменяет тон, и (2) инактивация мозжечкового выхода путем вливания мусцимола в межположительное ядро устраняет экспрессию CR, что позволяет предположить что мозжечковое обучение отвечает не только за то, нужно ли и когда реагировать, но и за то, насколько сильно реагировать.Эти данные показывают, что эта новая тренировочная процедура представляет собой управляемую экспериментальную парадигму для изучения мозжечковых механизмов, которые обучаются и кодируют амплитуду ответа.
Когда амплитуда ответа соответствует заранее заданной целевой амплитуде или превышает ее, УЗИ не выполняется (см.). Мы демонстрируем, что амплитуда ответа приближается к целевым амплитудам в диапазоне от 2 до 5 мм, со значительными групповыми различиями при анализе после 10 тренировок. Участие мозжечка в этой обучающей задаче подтверждается двумя наблюдениями: (1) амплитуда ответа может быть определена, когда стимуляция мшистых волокон заменяет тон, и (2) инактивация мозжечкового выхода путем вливания мусцимола в межположительное ядро устраняет экспрессию CR, что позволяет предположить что мозжечковое обучение отвечает не только за то, нужно ли и когда реагировать, но и за то, насколько сильно реагировать.Эти данные показывают, что эта новая тренировочная процедура представляет собой управляемую экспериментальную парадигму для изучения мозжечковых механизмов, которые обучаются и кодируют амплитуду ответа.
Схематическое изображение тренировочных процедур, используемых для тренировки реакции век на заданную амплитуду. Каждое тренировочное испытание включало предъявление тонального сигнала CS и запланированную доставку ультразвуковой стимуляции глаза с межстимульным интервалом 500 мс. Фактическая доставка УЗИ зависела от амплитуды ответа путем мониторинга амплитуды ответа в реальном времени во время CS.За десять миллисекунд до запланированной доставки США (время принятия решения в США) обучающее программное обеспечение было запрограммировано таким образом, чтобы определять, будут ли доставлены УЗИ: для ответов, которые соответствовали или превышали заданную амплитуду, доставлялись УЗИ; для ответов, которые не достигли целевой амплитуды, доставлялся УЗИ.
Каждое тренировочное испытание включало предъявление тонального сигнала CS и запланированную доставку ультразвуковой стимуляции глаза с межстимульным интервалом 500 мс. Фактическая доставка УЗИ зависела от амплитуды ответа путем мониторинга амплитуды ответа в реальном времени во время CS.За десять миллисекунд до запланированной доставки США (время принятия решения в США) обучающее программное обеспечение было запрограммировано таким образом, чтобы определять, будут ли доставлены УЗИ: для ответов, которые соответствовали или превышали заданную амплитуду, доставлялись УЗИ; для ответов, которые не достигли целевой амплитуды, доставлялся УЗИ.
Результаты
Получение целевой амплитуды отклика
В естественных условиях, таких как адаптация VOR, преследования или саккад, вызванное ошибкой возбуждающее воздействие на вход восходящего волокна в мозжечок уменьшается по мере того, как усиление отклика приближается к правильному значению (Гилберт и Тач, 1977; Раймонд и Лисбергер, 1997, 1998). Наша процедура обучения предназначена для более точного приближения к этим естественным условиям обучения путем связывания УЗ-презентации с амплитудой ответа относительно целевой амплитуды. Для этого мы тренировали четыре группы животных в течение 10 дней, каждая группа подвергалась одной из четырех целевых амплитуд (2, 3, 4 или 5 мм). Двадцать четыре кролика были случайным образом отнесены к одной из этих групп, при этом два животных из группы 5 мм в конечном итоге были исключены из-за технических трудностей. Для каждого ежедневного сеанса УЗИ не проводилось в испытаниях, в которых ответ век достигал целевой амплитуды в пределах 10 мс после введения УЗИ.УЗИ проводилось в испытаниях, в которых реакция века не достигала целевой амплитуды.
Наша процедура обучения предназначена для более точного приближения к этим естественным условиям обучения путем связывания УЗ-презентации с амплитудой ответа относительно целевой амплитуды. Для этого мы тренировали четыре группы животных в течение 10 дней, каждая группа подвергалась одной из четырех целевых амплитуд (2, 3, 4 или 5 мм). Двадцать четыре кролика были случайным образом отнесены к одной из этих групп, при этом два животных из группы 5 мм в конечном итоге были исключены из-за технических трудностей. Для каждого ежедневного сеанса УЗИ не проводилось в испытаниях, в которых ответ век достигал целевой амплитуды в пределах 10 мс после введения УЗИ.УЗИ проводилось в испытаниях, в которых реакция века не достигала целевой амплитуды.
Как показано на, животные в четырех группах приобрели устойчивые CR, и для каждой группы средняя амплитуда / величина ответов приближается к заданной цели. Для измерения амплитуды были включены все ответы, тогда как при измерении амплитуды были исключены все не-CR (те, у которых амплитуда <0,3 мм). Каждый из отдельных двухсторонних смешанных дисперсионных анализов (один для амплитуды и один для величины) показал значительную группу (амплитуда, F (3,198) = 7.43, p = 0,0039; величина, F = 11,18, p = 0,00013), тренировочный день (амплитуда, F (9,162) = 126,4, p <0,0001; величина, F = 101,21, p <0,0001 ) и взаимодействие группа × день (амплитуда, F (27,162) = 2,99, p <0,0001; величина, F = 3,66, p <0,0001). Эти анализы показывают, что животные учились, что между группами были различия в амплитуде и величине ответов, и что эти различия развивались в процессе обучения.Несмотря на эти различия в амплитуде и величине ответа, группы были сопоставимы по доле испытаний, в которых произошел полный ответ ( C ). Для этих данных двусторонний смешанный дисперсионный анализ показал значительный эффект дня тренировки ( F (9,162) = 123,8, p <0,0001), но не было никаких эффектов взаимодействия группы или группы × день тренировки ( p > 0,05).
Каждый из отдельных двухсторонних смешанных дисперсионных анализов (один для амплитуды и один для величины) показал значительную группу (амплитуда, F (3,198) = 7.43, p = 0,0039; величина, F = 11,18, p = 0,00013), тренировочный день (амплитуда, F (9,162) = 126,4, p <0,0001; величина, F = 101,21, p <0,0001 ) и взаимодействие группа × день (амплитуда, F (27,162) = 2,99, p <0,0001; величина, F = 3,66, p <0,0001). Эти анализы показывают, что животные учились, что между группами были различия в амплитуде и величине ответов, и что эти различия развивались в процессе обучения.Несмотря на эти различия в амплитуде и величине ответа, группы были сопоставимы по доле испытаний, в которых произошел полный ответ ( C ). Для этих данных двусторонний смешанный дисперсионный анализ показал значительный эффект дня тренировки ( F (9,162) = 123,8, p <0,0001), но не было никаких эффектов взаимодействия группы или группы × день тренировки ( p > 0,05).
Получение условных ответов век за 10 дней обучения для четырех групп, которым были назначены разные целевые амплитуды (2, 3, 4 и 5 мм). A , Амплитуда условных ответов век за 10 дней тренировки показывает получение разных амплитуд ответов для четырех групп. B , Аналогичные результаты были получены для измерения величины отклика, которая исключила ответы, которые не достигли критерия 0,3 мм, чтобы считаться CR. C , Поскольку вероятность ответа была устойчивой для всех четырех групп, результаты для амплитуды и величины были довольно схожими.В случаях, когда планки погрешностей (± SEM) не видны, ошибка меньше символа.
Есть два дополнительных индикатора, которые подтверждают гипотезу о том, что эта процедура ставит амплитуду ответа под экспериментальный контроль и более точно соответствует естественным условиям. Во-первых, должно быть уменьшение доли испытаний, в которых проводится УЗИ, с почти 1,0 до среднего уровня, что указывает на то, что амплитуда ответа иногда больше, а иногда меньше целевого. A показывает, что действительно доля испытаний, в которых выполнялась УЗИ, уменьшилась по сравнению с тренировкой, что указывает на увеличение количества испытаний, в которых целевая амплитуда была достигнута или превышена. Для этого показателя наблюдалось значительное влияние групп ( F (3,198) = 7,13, p = 0,0047) и тренировочного дня ( F (9,162) = 70,93, p <0,0001 ), но сравнение взаимодействий не было статистически надежным ( F (27,162) = 1.67, p = 0,0545). Во-вторых, должно происходить уменьшение средней ошибки, то есть среднего абсолютного значения разницы между амплитудой отклика и целевой амплитудой. Этот показатель отображается как функция количества тренировочных дней в B . На общее уменьшение средней ошибки указывает значительный эффект тренировочных дней ( F (9 162) = 102,17, p <0,0001). Поскольку начальное значение ошибки различается для четырех групп, неудивительно, что также наблюдался значительный групповой эффект ( F (3,198) = 14.
A показывает, что действительно доля испытаний, в которых выполнялась УЗИ, уменьшилась по сравнению с тренировкой, что указывает на увеличение количества испытаний, в которых целевая амплитуда была достигнута или превышена. Для этого показателя наблюдалось значительное влияние групп ( F (3,198) = 7,13, p = 0,0047) и тренировочного дня ( F (9,162) = 70,93, p <0,0001 ), но сравнение взаимодействий не было статистически надежным ( F (27,162) = 1.67, p = 0,0545). Во-вторых, должно происходить уменьшение средней ошибки, то есть среднего абсолютного значения разницы между амплитудой отклика и целевой амплитудой. Этот показатель отображается как функция количества тренировочных дней в B . На общее уменьшение средней ошибки указывает значительный эффект тренировочных дней ( F (9 162) = 102,17, p <0,0001). Поскольку начальное значение ошибки различается для четырех групп, неудивительно, что также наблюдался значительный групповой эффект ( F (3,198) = 14. 41, p <0,0001) и значительный эффект группы × тренировочные дни ( F (27,162) = 13,6, p <0,0001).
41, p <0,0001) и значительный эффект группы × тренировочные дни ( F (27,162) = 13,6, p <0,0001).
A , B , Уменьшение вероятности доставки в США ( A ) и средней ошибки ( B ) в зависимости от 10 дней обучения для четыре группы амплитуд (2, 3, 4 и 5 мм). Снижение вероятности УЗИ ( A ) указывает на увеличение доли ответов, которые соответствуют целевой амплитуде или превышают ее.Все четыре группы показали значительное снижение вероятности поражения в США, достигнув значений от 21 до 44%. B , Средняя ошибка уменьшилась до аналогичных значений во всех четырех группах.
В целом, эти анализы показывают, что (1) группы, разработанные с разницей в тренировках по амплитуде и величине ответа, (2) доля испытаний, в которых проводилось УЗИ, и средняя ошибка уменьшались за время обучения, и (3) там Нет различий в вероятности ответа для трех групп. Поскольку наиболее простой способ оценить свойства этого обучения – это оценить производительность в стабильном или близком к нему состоянии, далее мы сосредоточили анализ на данных за последние 2 дня обучения.
Поскольку наиболее простой способ оценить свойства этого обучения – это оценить производительность в стабильном или близком к нему состоянии, далее мы сосредоточили анализ на данных за последние 2 дня обучения.
Анализ асимптотической производительности
Для анализа стабильной производительности данные были объединены с 9-го и 10-го дней сбора данных для каждого субъекта. Этот анализ выявил четкие различия в амплитуде и величине CR между четырьмя группами ( A , B ).Отдельные односторонние дисперсионные анализы для амплитуды ( F (3,18) = 19,53, p <0,0001) и величины ( F = 37,25, p <0,0001) подтверждают надежность этих различий. Для измерения величины все шесть парных сравнений (критерий Тьюки, p <0,05) были надежными, а для амплитуды только сравнение 3 и 4 мм не было надежным. В отличие от этих различий, не было достоверных различий в измерении средней ошибки между четырьмя группами ( C ). В качестве финального теста на различия в амплитуде и величине ответов по четырем целевым амплитудным группам были рассчитаны нормализованные совокупные частотные распределения для каждой меры ответа ( G ). Тест Колмогорова – Смирнова ( p <0,005) показал, что все шесть парных сравнений для каждой группы были статистически надежными.
В качестве финального теста на различия в амплитуде и величине ответов по четырем целевым амплитудным группам были рассчитаны нормализованные совокупные частотные распределения для каждой меры ответа ( G ). Тест Колмогорова – Смирнова ( p <0,005) показал, что все шесть парных сравнений для каждой группы были статистически надежными.
Сравнения четырех амплитудных групп при стабильных показателях, взятых для каждого животного с 9-го и 10-го дней. A , Установившаяся амплитуда ответа различалась для четырех групп.Четыре пунктирные линии указывают целевую амплитуду для каждой группы. Все попарные сравнения (критерий Тьюки, p <0,05) были значимыми, за исключением сравнения между группами 3 и 4 мм. B , тот же формат, что и A для данных величины, которые представляют собой среднюю амплитуду всех CR без исключения не-CR. Все попарные сравнения были значимыми (критерий Тьюки, p <0,05). C , Стационарная средняя ошибка (абсолютное значение) существенно не различалась для четырех групп. D , Наблюдалась общая тенденция увеличения доли испытаний, в которых использовались УЗИ, с увеличением амплитуды цели. Единственное парное сравнение, которое было статистически надежным, было 2 против 5 мм. E , Все четыре группы показывают сопоставимые характеристики с точки зрения вероятности CR; между четырьмя группами не было значительных различий. F , Была общая тенденция к увеличению пиковой скорости CR по мере увеличения целевой амплитуды.Единственное достоверное (критерий Тьюки, p <0,05) сравнение было 2 против 5 мм. G , Суммарная доля ответов как функция амплитуды условного ответа (вверху) и величины (внизу) для четырех групп. Каждый CR для каждого животного помещали в бункер (ширина бункера 0,2 мм). Все шесть парных сравнений являются статистически надежными (критерий Колмогорова – Смирнова, p <0,005) для амплитуды, а также для измерения величины (кроме 4 против 5 мм, p <0.
C , Стационарная средняя ошибка (абсолютное значение) существенно не различалась для четырех групп. D , Наблюдалась общая тенденция увеличения доли испытаний, в которых использовались УЗИ, с увеличением амплитуды цели. Единственное парное сравнение, которое было статистически надежным, было 2 против 5 мм. E , Все четыре группы показывают сопоставимые характеристики с точки зрения вероятности CR; между четырьмя группами не было значительных различий. F , Была общая тенденция к увеличению пиковой скорости CR по мере увеличения целевой амплитуды.Единственное достоверное (критерий Тьюки, p <0,05) сравнение было 2 против 5 мм. G , Суммарная доля ответов как функция амплитуды условного ответа (вверху) и величины (внизу) для четырех групп. Каждый CR для каждого животного помещали в бункер (ширина бункера 0,2 мм). Все шесть парных сравнений являются статистически надежными (критерий Колмогорова – Смирнова, p <0,005) для амплитуды, а также для измерения величины (кроме 4 против 5 мм, p <0. 05).
05).
A – C в совокупности предполагают, что, хотя средняя ошибка амплитуды была сопоставима в четырех группах, источники ошибок были разными. Ответы для групп 2 и 3 мм имели тенденцию выходить за цель, тогда как ответы для группы 5 мм имели тенденцию выходить за пределы цели. Эту тенденцию можно вывести из сравнения вероятности в США, показанного в D , которое отражает долю ответов, которые не достигли целевого значения. Несмотря на четкую тенденцию к увеличению этого показателя с целевой амплитудой, различия между группами только приближались к значимости ( F (3,18) = 3.44, p = 0,08). Только группа 2 мм показала значительную тенденцию к превышению целевого значения, потому что отдельные тесты t для каждой группы выявили только достоверное отличие от среднего значения популяции в 50% (равные реакции недостижения и превышения) для группы 2 мм ( t ). (5) = 3,47, p <0,05). Несмотря на разницу в амплитуде и величине ответа, группы показали сопоставимые показатели ( F (3,18) = 1,799, p = 0. 37) с точки зрения вероятности CR ( E ). Наконец, F показывает, что существует также общая тенденция увеличения максимальной скорости КЛ с целевой амплитудой ( F (3,18) = 7,81, p = 0,003). Парные сравнения показали, что различия 2 по сравнению с 4 мм и 2 по сравнению с 5 мм были надежными (критерий Тьюки, p <0,05).
37) с точки зрения вероятности CR ( E ). Наконец, F показывает, что существует также общая тенденция увеличения максимальной скорости КЛ с целевой амплитудой ( F (3,18) = 7,81, p = 0,003). Парные сравнения показали, что различия 2 по сравнению с 4 мм и 2 по сравнению с 5 мм были надежными (критерий Тьюки, p <0,05).
Предыдущие исследования показали, что точное время CR является характерной чертой кондиционирования век (Smith, 1968; Smith et al., 1969; Милленсон и др., 1977; Маук и Руис, 1992). Время ожидания и время нарастания ответов систематически меняются в зависимости от межстимульного интервала, так что ответы имеют тенденцию достигать пика около времени доставки УЗИ. Мы исследовали различные меры времени ответа для четырех целевых амплитудных групп и не обнаружили достоверных различий во времени ответа ( A ). Ни время ожидания начала, ни время ожидания критерия, время ожидания максимальной скорости, ни время ожидания ответа пика не показали статистически достоверных различий между группами.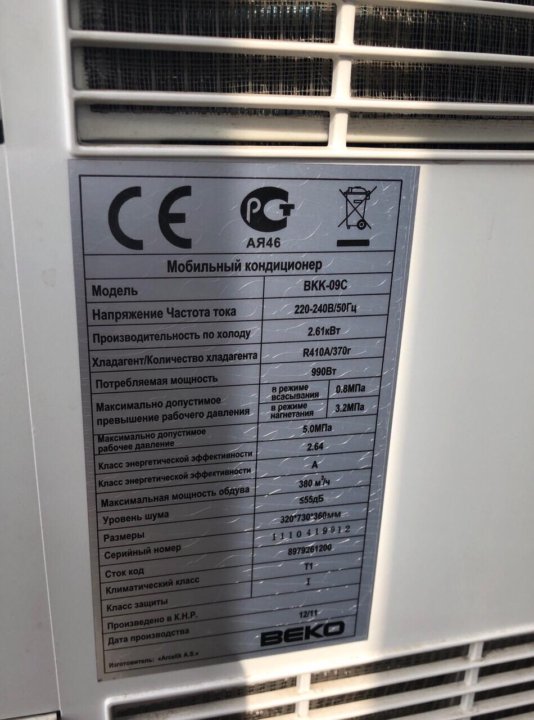 Когда ответы были разделены в зависимости от того, превышают ли они цель (испытания в США) или ниже цели (испытания в США), наблюдались значительные различия во времени ответов во всех четырех группах ( B ). Отдельные двухсторонние смешанные ANOVA для задержки начала и задержки до максимальной скорости показали только значимое влияние типа ответа (не в США по сравнению с США; начало, F (1,18) = 100,6, p < 0,0001; пиковая скорость, F = 49.04, p <0,0001). Этот анализ показывает, что в целом ответы, которые соответствовали целевой амплитуде или превышали ее, имели более короткие задержки, чем ответы, которые не соответствовали целевой амплитуде.
Когда ответы были разделены в зависимости от того, превышают ли они цель (испытания в США) или ниже цели (испытания в США), наблюдались значительные различия во времени ответов во всех четырех группах ( B ). Отдельные двухсторонние смешанные ANOVA для задержки начала и задержки до максимальной скорости показали только значимое влияние типа ответа (не в США по сравнению с США; начало, F (1,18) = 100,6, p < 0,0001; пиковая скорость, F = 49.04, p <0,0001). Этот анализ показывает, что в целом ответы, которые соответствовали целевой амплитуде или превышали ее, имели более короткие задержки, чем ответы, которые не соответствовали целевой амплитуде.
Меры времени отклика по сравнению с четырьмя группами амплитуды. A , Четыре графика показывают задержку до начала, задержку до критерия, задержку до пиковой скорости и задержку до пиковой амплитуды (для испытаний за пределами США). Статистически достоверной разницы между группами по этим показателям ответа не было. B , Разница во времени испытаний в США и за пределами США. Раздельный анализ исследований в УЗИ (амплитуда ниже целевого) и неамериканских исследований (амплитуда на уровне или выше) выявил общую тенденцию к более быстрым и коротким задержкам ответов в неамериканских исследованиях. Все восемь сравнений (4 группы × 2 меры ответа) были статистически надежными (парный тест t , p <0,01).
B , Разница во времени испытаний в США и за пределами США. Раздельный анализ исследований в УЗИ (амплитуда ниже целевого) и неамериканских исследований (амплитуда на уровне или выше) выявил общую тенденцию к более быстрым и коротким задержкам ответов в неамериканских исследованиях. Все восемь сравнений (4 группы × 2 меры ответа) были статистически надежными (парный тест t , p <0,01).
Примеры необработанных ответов от одного испытуемого из каждой амплитудной группы (день 10 в каждом случае) показаны в.Для каждого субъекта ответы, которые соответствовали целевой амплитуде или превышали ее, показаны в левом столбце, а те, которые не соответствовали целевой амплитуде (и, следовательно, были доставлены УЗИ), показаны в правом столбце. Эти данные показывают общую тенденцию реакции к перерегулированию для меньшей амплитуды цели и к недолету для цели 5 мм. Вместе эти анализы демонстрируют, что процедура тренировки целевой амплитуды может использоваться для получения разумного экспериментального контроля над амплитудой ответа при кондиционировании век, что средняя ошибка амплитуды сопоставима для целей от 2 до 5 мм, и что эти различия возникают на сопоставимых уровнях CR. и сопоставимые статистические данные о сроках проведения CR.
и сопоставимые статистические данные о сроках проведения CR.
Пример необработанных ответов от четырех животных, по одному от каждой группы амплитуд, взятых с 10-го дня обучения. Каждая развертка представляет собой отдельную реакцию века (первая проба впереди), причем черная область каждой развертки указывает продолжительность CS. Ответы были разделены на испытания, проводимые за пределами США (левый столбец) и испытания в США (правый столбец). Последовательность испытаний каждого животного в США и за пределами США очевидна по интервалу между испытаниями. Обратите внимание на общую тенденцию к большему количеству испытаний без УЗИ с малыми амплитудами цели и к большему количеству испытаний УЗИ с большими амплитудами цели.Компонент рефлекторного ответа каждого испытания виден в светло-серой области испытаний «With-US».
Амплитуда отклика после повторного обнаружения новой цели
Практическая и концептуальная важность тренировки амплитуды целевой будет уменьшена, если амплитуды отклика цели не могут быть получены и повторно получены в соответствии с планом эксперимента. Поэтому мы тестировали каждое животное, используя новую целевую амплитуду, отличную от начальной. После 10 дней обучения до начальной целевой амплитуды каждый субъект был обучен в течение 10 дополнительных дней с использованием новой целевой амплитуды.Новые целевые амплитуды для групп 2, 3, 4 и 5 мм, соответственно, составляли 4, 5, 2 и 3 мм. показывает сравнение амплитуд ответа для этих четырех групп для начальной и новой целевой амплитуд. Как и выше, эти данные включали объединенное среднее значение 9 и 10 дней для каждого субъекта. Мы начали с того, что спросили, есть ли различия между получением и повторным обнаружением для каждой из четырех целевых амплитуд (левый график). Это сравнение между субъектами сравнивает каждую цель, когда она использовалась при захвате, по сравнению с повторным захватом (т.е., получение от группы 2 мм и повторное обнаружение на цели 2 мм, первоначально группы 4 мм). Двусторонний дисперсионный анализ выявил только основной эффект целевой амплитуды ( F (3,36) = 37,06, p <0,0001), эффекты захвата по сравнению с повторным захватом и взаимодействия не были значительными, и не было надежных различия в амплитудах, полученных во время захвата, по сравнению с повторным обнаружением новой цели.
Поэтому мы тестировали каждое животное, используя новую целевую амплитуду, отличную от начальной. После 10 дней обучения до начальной целевой амплитуды каждый субъект был обучен в течение 10 дополнительных дней с использованием новой целевой амплитуды.Новые целевые амплитуды для групп 2, 3, 4 и 5 мм, соответственно, составляли 4, 5, 2 и 3 мм. показывает сравнение амплитуд ответа для этих четырех групп для начальной и новой целевой амплитуд. Как и выше, эти данные включали объединенное среднее значение 9 и 10 дней для каждого субъекта. Мы начали с того, что спросили, есть ли различия между получением и повторным обнаружением для каждой из четырех целевых амплитуд (левый график). Это сравнение между субъектами сравнивает каждую цель, когда она использовалась при захвате, по сравнению с повторным захватом (т.е., получение от группы 2 мм и повторное обнаружение на цели 2 мм, первоначально группы 4 мм). Двусторонний дисперсионный анализ выявил только основной эффект целевой амплитуды ( F (3,36) = 37,06, p <0,0001), эффекты захвата по сравнению с повторным захватом и взаимодействия не были значительными, и не было надежных различия в амплитудах, полученных во время захвата, по сравнению с повторным обнаружением новой цели. Правая часть показывает те же данные, построенные заново, чтобы выделить сравнения амплитуд, полученных для сбора и повторного сбора данных в одних и тех же группах.Четыре отдельных парных теста t ( p <0,01) показали, что для каждой группы амплитуда ответов, полученных при обнаружении одной цели, отличалась от тех, которые были получены при повторном обнаружении на новую цель. Эти анализы показывают, что как получение первой целевой амплитуды, так и повторное обнаружение новой целевой амплитуды вызвали значительную разницу в амплитуде ответа для каждой целевой амплитудной группы и что не было никаких различий в этих тенденциях между захватом и повторным захватом.
Правая часть показывает те же данные, построенные заново, чтобы выделить сравнения амплитуд, полученных для сбора и повторного сбора данных в одних и тех же группах.Четыре отдельных парных теста t ( p <0,01) показали, что для каждой группы амплитуда ответов, полученных при обнаружении одной цели, отличалась от тех, которые были получены при повторном обнаружении на новую цель. Эти анализы показывают, что как получение первой целевой амплитуды, так и повторное обнаружение новой целевой амплитуды вызвали значительную разницу в амплитуде ответа для каждой целевой амплитудной группы и что не было никаких различий в этих тенденциях между захватом и повторным захватом.
Сравнение устойчивых амплитуд ответа после первоначального сбора данных (темно-серый; те же данные, что и A ) и у тех же животных после 10 дней повторного обнаружения с новой целевой амплитудой. Начальная амплитуда цели и амплитуда цели повторного обнаружения были, соответственно, для четырех групп (от 2 до 4 мм, от 3 до 5 мм, от 4 до 2 мм и от 5 до 3 мм). Как и в случае, данные получены из среднего значения для каждого животного за 9-й и 10-й дни захвата (и повторного захвата). Слева: сравнение идентичных целевых амплитуд для захвата и повторного захвата.Статистически достоверных различий для каждой целевой амплитуды между данными сбора и повторного обнаружения не было ( t тестов). Целевая амплитуда обозначена пунктирными линиями. Справа: сравнение данных приобретения и повторного приобретения для одних и тех же групп животных. Различные целевые амплитуды захвата и повторного захвата для каждой группы снова обозначены пунктирными линиями. Парные тесты t ( p <0,01) показали статистически достоверные различия между амплитудами сбора и повторного обнаружения для каждой группы.
Как и в случае, данные получены из среднего значения для каждого животного за 9-й и 10-й дни захвата (и повторного захвата). Слева: сравнение идентичных целевых амплитуд для захвата и повторного захвата.Статистически достоверных различий для каждой целевой амплитуды между данными сбора и повторного обнаружения не было ( t тестов). Целевая амплитуда обозначена пунктирными линиями. Справа: сравнение данных приобретения и повторного приобретения для одних и тех же групп животных. Различные целевые амплитуды захвата и повторного захвата для каждой группы снова обозначены пунктирными линиями. Парные тесты t ( p <0,01) показали статистически достоверные различия между амплитудами сбора и повторного обнаружения для каждой группы.
Стимуляция мшистых волокон в качестве CS
Значительные доказательства указывают на то, что приобретение и экспрессия CR во время формирования павлового (отложенного) века опосредуется пластичностью мозжечка. Поскольку обусловленность целевой амплитуды, строго говоря, не является павловской, мы провели два дополнительных эксперимента, чтобы проверить возможность того, что обучение в этих условиях опосредуется нецеребеллярными механизмами. Способность заменять стимуляцию мшистых волокон тонусом CS – одно из наиболее убедительных доказательств, подтверждающих, что мозжечковые механизмы являются основой кондиционирования век.Таким образом, мы сравнили эффекты тренировки с целевой амплитудой 3 мм, используя тон CS в одной группе и стимуляцию мшистых волокон в качестве CS во второй группе. В остальном процедуры были идентичны процедурам, использованным выше для группы 3 мм. показывает результаты за 10 дней тренировок для этих двух групп. A – D сравнивает эти две группы за 10 дней обучения, а E показывает сравнения в установившемся состоянии, как в предыдущем эксперименте. Отдельные двухфакторные смешанные ANOVA для четырех мер реакции показали только основные эффекты тренировочных дней (амплитуда, F (9,54) = 30.54; ошибка, F = 17,71; Вероятность CR, F = 29,09; Вероятность США – 16,62; p <0,0001 во всех случаях). Ни один из основных эффектов группы или эффектов взаимодействия не был значимым, равно как и сравнение тона и мшистого волокна ( E ) для восьми показателей реакции, которые были проанализированы для данных устойчивого состояния (дни 9 и 10, t тесты, p > 0,05).
Способность заменять стимуляцию мшистых волокон тонусом CS – одно из наиболее убедительных доказательств, подтверждающих, что мозжечковые механизмы являются основой кондиционирования век.Таким образом, мы сравнили эффекты тренировки с целевой амплитудой 3 мм, используя тон CS в одной группе и стимуляцию мшистых волокон в качестве CS во второй группе. В остальном процедуры были идентичны процедурам, использованным выше для группы 3 мм. показывает результаты за 10 дней тренировок для этих двух групп. A – D сравнивает эти две группы за 10 дней обучения, а E показывает сравнения в установившемся состоянии, как в предыдущем эксперименте. Отдельные двухфакторные смешанные ANOVA для четырех мер реакции показали только основные эффекты тренировочных дней (амплитуда, F (9,54) = 30.54; ошибка, F = 17,71; Вероятность CR, F = 29,09; Вероятность США – 16,62; p <0,0001 во всех случаях). Ни один из основных эффектов группы или эффектов взаимодействия не был значимым, равно как и сравнение тона и мшистого волокна ( E ) для восьми показателей реакции, которые были проанализированы для данных устойчивого состояния (дни 9 и 10, t тесты, p > 0,05). Эти данные исключают факторы, расположенные выше мозжечка, как способствующие приобретению амплитуды ответа, и подтверждают гипотезу о том, что кондиционирование век к целевой амплитуде опосредуется мозжечком.
Эти данные исключают факторы, расположенные выше мозжечка, как способствующие приобретению амплитуды ответа, и подтверждают гипотезу о том, что кондиционирование век к целевой амплитуде опосредуется мозжечком.
Получение целевой амплитуды (3 мм) для животных, обученных с помощью тонального CS и стимуляции мшистыми волокнами в качестве CS. A , Амплитуда CR за 10 дней обучения. B , Средняя ошибка (абсолютное значение) за 10 дней обучения. MF, Мшистое волокно. C , D , вероятность CR ( C ) и вероятность США ( D ) за 10 дней обучения. E , Сравнение между группами CS тональной стимуляции и CS со стимуляцией мшистыми волокнами для различных мер реакции, взятых с 9-го и 10-го дней тренировки.Ни одно из шести сравнений групп не было статистически надежным (тест t , p > 0,05).
Обратимая инактивация мозжечка
В качестве дополнительной проверки гипотезы о том, что мозжечок необходим для кондиционирования до заданной амплитуды, экспрессия CR была протестирована на четырех животных, обученных тону, и четырех животных, обученных мшистым волокнам, до и после инактивации. мозжечкового выхода через инфузию мусцимола, агониста ГАМК, в переднее межположительное ядро ().Субъекты в обеих группах тестировались в отдельные дни с инфузиями носителя или инфузиями мусцимола (порядок уравновешивался для каждой группы). График средних показателей по группе (в центре) показывает, что для обеих групп не было достоверных изменений после инфузии носителя, тогда как инфузии мусцимола отменяли экспрессию CR. Необработанные данные для одного субъекта стимуляции мшистых волокон для инфузий носителя (слева) и мусцимола (справа) показаны вверху. Пример канюли (слева) и стимулирующих электродов от того же пациента показаны внизу.
Необходимость мозжечка для выражения приобретенных ответов на заданную амплитуду 3 мм. Две группы, одна из которых использовала тональный CS, а другая, в которой стимуляция волокон использовалась в качестве CS, были протестированы в отдельные дни либо с инфузией носителя, либо с инфузией мусцимола, агониста ГАМК. На двух графиках показаны средние показатели по группе отсутствия эффекта от инфузии носителя (слева) и отмены CR с помощью инфузии мусцимола (справа). Время настаивания в каждом случае указано черной стрелкой.Примеры исходных данных от одного животного показаны выше: следы слева – это инфузия носителя, а следы справа – инфузия мусцимола. В каждом случае ответы до инфузии находятся впереди (первое испытание впереди), а реакции после инфузии – сзади. На двух нижних фотографиях показано размещение инфузионной канюли (слева) и стимулирующих электродов из мшистых волокон (справа) для животного, исходные данные которого показаны выше. Этому животному были имплантированы два электрода из покрытых мхом волокон; только один был использован для этого эксперимента.MF, Мшистые волокна.
В совокупности исследования стимуляции мшистых волокон и инактивации глубоких ядер показывают (1), что мозжечок необходим для экспрессии CR, продуцируемых кондиционированием века до целевой амплитуды (результаты инфузии), и (2) что механизмы, ответственные за приобретение и экспрессия целевой амплитуды ответа не находятся в пути CR выше мозжечка (результаты стимуляции мшистыми волокнами).
Обсуждение
Мы адаптировали традиционные процедуры кондиционирования павловских век, чтобы ввод ошибки, определяемый США, зависел от амплитуды CR.Пропустив УЗИ в исследованиях, в которых CR соответствует или превышает заданную целевую амплитуду (и представив УЗИ в противном случае), мы показали, что результирующие условные реакции век имеют тенденцию к целевой амплитуде. Это верно как для получения исходной целевой амплитуды, так и для повторного обнаружения с использованием другой цели. Несмотря на то, что наблюдалась тенденция к перерегулированию при малых амплитудах цели и к недолету с наибольшей целью, средняя абсолютная ошибка не различалась для групп, обученных для целевых амплитуд от 2 до 5 мм. Эти данные показывают, что, адаптируя процедуры кондиционирования век к естественным условиям обучения мозжечка, амплитуду CR можно поставить под экспериментальный контроль. Способность устранять эти CR с помощью инактивации мозжечка и способность заменять стимуляцию мшистых волокон тональным CS подтверждают гипотезу о том, что, как и в случае кондиционирования века в форме павлова, кондиционирование века до целевой амплитуды опосредуется мозжечком. Эти результаты формируют основу для систематических и контролируемых исследований того, как мозжечок обучается, сохраняет и выражает надлежащую амплитуду откликов, которые он адаптирует.
Эти данные показывают, что, адаптируя процедуры кондиционирования век к естественным условиям обучения мозжечка, амплитуду CR можно поставить под экспериментальный контроль. Способность устранять эти CR с помощью инактивации мозжечка и способность заменять стимуляцию мшистых волокон тональным CS подтверждают гипотезу о том, что, как и в случае кондиционирования века в форме павлова, кондиционирование века до целевой амплитуды опосредуется мозжечком. Эти результаты формируют основу для систематических и контролируемых исследований того, как мозжечок обучается, сохраняет и выражает надлежащую амплитуду откликов, которые он адаптирует.
Значение для кондиционирования век и мозжечка
Необходимость мозжечка для кондиционирования века с задержкой павлова подтверждается большим количеством данных. Настоящие результаты показывают, что непредвиденные обстоятельства Павлова, при которых доставка УЗИ не зависит от CR, не являются особенностью, которая делает мозжечок необходимым. Поскольку, строго говоря, процедуры тренировки до целевой амплитуды включают в себя оперантные непредвиденные обстоятельства, необходимость мозжечка в кондиционировании век связана с природой стимулов и системой реакции, а не с природой непредвиденных обстоятельств, которые управляют представлением усиливающий стимул.
Настоящие данные также обращаются к распространенному заблуждению о кондиционировании век, а именно, что это рефлекторное кондиционирование или кондиционирование моргания глаз. Павлов выступил с идеей, что классическое кондиционирование включает в себя пути, активируемые CS, обретающие способность активировать путь, который опосредует рефлекторный ответ (Павлов, 1927). Нервные пути, которые, как известно, опосредуют экспрессию CR и рефлекторные мигания, вызываемые США, не согласуются с гипотезой Павлова. Рефлекторные мигания, вызываемые первичными афферентами, проходящими через верхний глазничный нерв, опосредуются двумя путями (Pellegrini et al., 1995). Первичные афференты проецируются как на каудально-межполярную пограничную область спинномозгового ядра тройничного нерва, так и на дорсальный рог, сегмент С1 спинного мозга. Обе эти области проецируются в область orbicularis oculi лицевого ядра. Эксперименты с поражением показали, что спинальный тройничный путь опосредует начальный компонент R1 рефлекторных морганий, а путь спинного мозга опосредует более длинный латентный компонент R2 (Pelligrini et al. , 1995). Путь CR, выступающий из переднего межположительного ядра через красное ядро, по-видимому, соединяется с этим путем в двигательных или премоторных нейронах лицевого ядра.Следовательно, рефлекторные мигания и CRs опосредуются совершенно разными нервными путями и, как можно ожидать, будут проявлять совершенно разные свойства, как было четко установлено в исследованиях на людях (Schade Powers et al., 2010). Поэтому условные реакции век более правильно охарактеризовать как «реакции век», чем «моргания глаз», и настоящие данные демонстрируют, что амплитуда этих ответов век, как и большинство других реакций скелетных мышц, может быть изучена и опосредована обучением мозжечка.
, 1995). Путь CR, выступающий из переднего межположительного ядра через красное ядро, по-видимому, соединяется с этим путем в двигательных или премоторных нейронах лицевого ядра.Следовательно, рефлекторные мигания и CRs опосредуются совершенно разными нервными путями и, как можно ожидать, будут проявлять совершенно разные свойства, как было четко установлено в исследованиях на людях (Schade Powers et al., 2010). Поэтому условные реакции век более правильно охарактеризовать как «реакции век», чем «моргания глаз», и настоящие данные демонстрируют, что амплитуда этих ответов век, как и большинство других реакций скелетных мышц, может быть изучена и опосредована обучением мозжечка.
Значение для обучения мозжечка усилению или амплитуде
Вопрос о том, как и где мозжечок обучается и сохраняет усиление или амплитуду ответа, взаимосвязан с вопросом о месте (ах) пластичности, ответственных за обучение мозжечка. На этот вопрос лучше всего ответить с помощью исследований, включающих анализ адаптации VOR (Raymond et al. , 1996; Boyden et al., 2004). Первоначальные усилия в этой области вызвали серьезные дискуссии о том, происходит ли пластичность в коре мозжечка или в вестибулярном ядре, чьи связи и схема параллельны глубоким ядрам мозжечка (Lisberger and Sejnowski, 1992, 1993; Ito, 1993).Более поздние теории предполагают участие пластичности как в коре мозжечка, так и в глубоком ядре (Perrett et al., 1993; Raymond et al., 1996; Mauk and Donegan, 1997). Когда задействовано более одного сайта пластичности, становится важным вопрос об относительном вкладе этих двух сайтов. Относительный вклад коры мозжечка и механизмов пластичности глубокого ядра мозжечка в усиление или амплитуду ответа остается нерешенным, и есть указания на то, что относительный вклад может варьироваться сложным образом в зависимости от обстоятельств (Mauk, 1997).Кондиционирование век до целевой амплитуды обеспечивает способ решения этих вопросов с помощью относительно простой и экспериментально управляемой системы, которая поддается параметрическому поведенческому анализу, обратимым повреждениям, in vivo записи и даже крупномасштабному компьютерному моделированию.
, 1996; Boyden et al., 2004). Первоначальные усилия в этой области вызвали серьезные дискуссии о том, происходит ли пластичность в коре мозжечка или в вестибулярном ядре, чьи связи и схема параллельны глубоким ядрам мозжечка (Lisberger and Sejnowski, 1992, 1993; Ito, 1993).Более поздние теории предполагают участие пластичности как в коре мозжечка, так и в глубоком ядре (Perrett et al., 1993; Raymond et al., 1996; Mauk and Donegan, 1997). Когда задействовано более одного сайта пластичности, становится важным вопрос об относительном вкладе этих двух сайтов. Относительный вклад коры мозжечка и механизмов пластичности глубокого ядра мозжечка в усиление или амплитуду ответа остается нерешенным, и есть указания на то, что относительный вклад может варьироваться сложным образом в зависимости от обстоятельств (Mauk, 1997).Кондиционирование век до целевой амплитуды обеспечивает способ решения этих вопросов с помощью относительно простой и экспериментально управляемой системы, которая поддается параметрическому поведенческому анализу, обратимым повреждениям, in vivo записи и даже крупномасштабному компьютерному моделированию.
Добавляя, сколько и когда
Abstract
Концептуальные и практические преимущества кондиционирования павловых век облегчают анализ вычислений и обучения мозжечка.Тем не менее, процедуры кондиционирования век во многом нереальны. Сигнал ошибки для оливоцеребеллярной системы не уменьшается, поскольку обучение адаптирует амплитуду или усиление ответа. Это по своей сути ограничивает полезность кондиционирования век для исследований, изучающих, как механизмы обучения мозжечка приобретают и сохраняют амплитуду адаптивного ответа. Мы сообщаем о разработке и описании процедуры обучения, в которой амплитуда условной реакции ставится под экспериментальный контроль с непредвиденными обстоятельствами, которые более близки к естественным условиям.В этой процедуре доставка безусловного стимула (US) зависит от амплитуды условного ответа: US доставляется для ответов, которые не достигают заданной целевой амплитуды, и опускается для ответов, которые соответствуют или превышают цель. Мы обнаружили, что кролики, обученные либо тональной стимуляции, либо стимуляции мшистыми волокнами в качестве условного стимула, усваивают ответы, которые достигают целевых амплитуд в диапазоне от 2 до 5 мм. Инактивация межположительного ядра с помощью инфузий мусцимола устраняет эти условные реакции, указывая на то, что участие мозжечка в кондиционировании век не связано явно с использованием процедур Павлова.Вместе с предыдущими исследованиями эти данные предполагают, что амплитуда ответа изучается и кодируется в мозжечке во время кондиционирования век. Таким образом, эти результаты обеспечивают основу для систематических и контролируемых исследований мозжечковых механизмов, которые обучаются и кодируют правильную амплитуду адаптивных движений.
Мы обнаружили, что кролики, обученные либо тональной стимуляции, либо стимуляции мшистыми волокнами в качестве условного стимула, усваивают ответы, которые достигают целевых амплитуд в диапазоне от 2 до 5 мм. Инактивация межположительного ядра с помощью инфузий мусцимола устраняет эти условные реакции, указывая на то, что участие мозжечка в кондиционировании век не связано явно с использованием процедур Павлова.Вместе с предыдущими исследованиями эти данные предполагают, что амплитуда ответа изучается и кодируется в мозжечке во время кондиционирования век. Таким образом, эти результаты обеспечивают основу для систематических и контролируемых исследований мозжечковых механизмов, которые обучаются и кодируют правильную амплитуду адаптивных движений.
Введение
Павловское кондиционирование века облегчило анализ обучения и вычислений мозжечка (McCormick and Thompson, 1984; Perrett et al., 1993; Маук и Донеган, 1997; Медина и Маук, 1999; Hansel et al., 2001; Medina et al., 2001, 2002), в основном из-за тесного соответствия между тренировочными стимулами и входными сигналами мозжечка, а также между выходной мощностью мозжечка и поведенческими реакциями (McCormick and Thompson, 1984; Mauk et al. , 1986; Steinmetz et al. , 1986; Mauk, Donegan, 1997; Hesslow et al., 1999). Идея Павлова (1927) о том, что условное кондиционирование – это «рефлексивное обучение», при котором условный стимул (КС) приобретает способность активировать рефлекторный путь, запускаемый подкрепляющим стимулом, является неправильным представлением, обычно применяемым к кондиционированию век.В самом деле, неправильное название «кондиционирование моргания», по-видимому, происходит из этого заблуждения. Данные, противоречащие этой идее, включают известные схемы кондиционирования век, которые ясно показывают, что путь CS не связан с рефлекторным путем через обучение (Mauk and Donegan, 1997; Thompson et al., 1998), из доказательств того, что условные реакции век не являются морганиями (Schade Powers et al., 2010), а из простого поведенческого наблюдения, что во время приобретения амплитуда условной реакции не является однозначной.С тренировкой амплитуда условной реакции монотонно растет от едва заметного до максимального закрытия век.
, 1986; Steinmetz et al. , 1986; Mauk, Donegan, 1997; Hesslow et al., 1999). Идея Павлова (1927) о том, что условное кондиционирование – это «рефлексивное обучение», при котором условный стимул (КС) приобретает способность активировать рефлекторный путь, запускаемый подкрепляющим стимулом, является неправильным представлением, обычно применяемым к кондиционированию век.В самом деле, неправильное название «кондиционирование моргания», по-видимому, происходит из этого заблуждения. Данные, противоречащие этой идее, включают известные схемы кондиционирования век, которые ясно показывают, что путь CS не связан с рефлекторным путем через обучение (Mauk and Donegan, 1997; Thompson et al., 1998), из доказательств того, что условные реакции век не являются морганиями (Schade Powers et al., 2010), а из простого поведенческого наблюдения, что во время приобретения амплитуда условной реакции не является однозначной.С тренировкой амплитуда условной реакции монотонно растет от едва заметного до максимального закрытия век.
Тем не менее, стандартные тренировочные процедуры кондиционирования век не позволяют экспериментально контролировать амплитуду ответа. Для более естественных реакций, таких как адаптация вестибулоокулярного рефлекса (VOR), вход ошибки в оливо-церебеллярную систему (т. Е. Входной сигнал восходящего волокна в мозжечок) уменьшается по мере того, как обучение адаптирует усиление или амплитуду ответа (Gilbert and Thach, 1977). ; Раймонд и Лисбергер, 1997, 1998).Поскольку вероятность сигнала ошибки в оливоцеребеллярной системе [через предъявление безусловного стимула (УЗ)] не уменьшается по мере роста амплитуды условного ответа, процедуры Павлова по своей природе способствуют условным ответам (КО) с максимальной амплитудой (5–6 мм для кроликов). ). Даже при воздушной затяжке, такой как УЗИ, CR (расширение мигательной перепонки) не может полностью устранить активацию УЗ-лазейных волокон, потому что внешние веки обычно ограничены, а сама мигательная перепонка весьма чувствительна.По этим причинам кондиционирование век не было полезным для анализа мозжечковых механизмов, участвующих в обучении и кодировании амплитуды ответа (то есть усиления ответа).
Мы сообщаем о разработке и описании процедуры тренировки век, которая более точно имитирует естественные требования к обучению мозжечка и ставит амплитуду ответа под экспериментальный контроль. В этой процедуре УЗИ зависит от амплитуды условного ответа, так же как ошибочные воздействия на мозжечок зависят от усиления ответа.Когда амплитуда ответа соответствует заранее заданной целевой амплитуде или превышает ее, УЗИ не выполняется (см.). Мы демонстрируем, что амплитуда ответа приближается к целевым амплитудам в диапазоне от 2 до 5 мм, со значительными групповыми различиями при анализе после 10 тренировок. Участие мозжечка в этой обучающей задаче подтверждается двумя наблюдениями: (1) амплитуда ответа может быть определена, когда стимуляция мшистых волокон заменяет тон, и (2) инактивация мозжечкового выхода путем вливания мусцимола в межположительное ядро устраняет экспрессию CR, что позволяет предположить что мозжечковое обучение отвечает не только за то, нужно ли и когда реагировать, но и за то, насколько сильно реагировать. Эти данные показывают, что эта новая тренировочная процедура представляет собой управляемую экспериментальную парадигму для изучения мозжечковых механизмов, которые обучаются и кодируют амплитуду ответа.
Эти данные показывают, что эта новая тренировочная процедура представляет собой управляемую экспериментальную парадигму для изучения мозжечковых механизмов, которые обучаются и кодируют амплитуду ответа.
Схематическое изображение тренировочных процедур, используемых для тренировки реакции век на заданную амплитуду. Каждое тренировочное испытание включало предъявление тонального сигнала CS и запланированную доставку ультразвуковой стимуляции глаза с межстимульным интервалом 500 мс. Фактическая доставка УЗИ зависела от амплитуды ответа путем мониторинга амплитуды ответа в реальном времени во время CS.За десять миллисекунд до запланированной доставки США (время принятия решения в США) обучающее программное обеспечение было запрограммировано таким образом, чтобы определять, будут ли доставлены УЗИ: для ответов, которые соответствовали или превышали заданную амплитуду, доставлялись УЗИ; для ответов, которые не достигли целевой амплитуды, доставлялся УЗИ.
Результаты
Получение целевой амплитуды отклика
В естественных условиях, таких как адаптация VOR, преследования или саккад, вызванное ошибкой возбуждающее воздействие на вход восходящего волокна в мозжечок уменьшается по мере того, как усиление отклика приближается к правильному значению (Гилберт и Тач, 1977; Раймонд и Лисбергер, 1997, 1998). Наша процедура обучения предназначена для более точного приближения к этим естественным условиям обучения путем связывания УЗ-презентации с амплитудой ответа относительно целевой амплитуды. Для этого мы тренировали четыре группы животных в течение 10 дней, каждая группа подвергалась одной из четырех целевых амплитуд (2, 3, 4 или 5 мм). Двадцать четыре кролика были случайным образом отнесены к одной из этих групп, при этом два животных из группы 5 мм в конечном итоге были исключены из-за технических трудностей. Для каждого ежедневного сеанса УЗИ не проводилось в испытаниях, в которых ответ век достигал целевой амплитуды в пределах 10 мс после введения УЗИ.УЗИ проводилось в испытаниях, в которых реакция века не достигала целевой амплитуды.
Наша процедура обучения предназначена для более точного приближения к этим естественным условиям обучения путем связывания УЗ-презентации с амплитудой ответа относительно целевой амплитуды. Для этого мы тренировали четыре группы животных в течение 10 дней, каждая группа подвергалась одной из четырех целевых амплитуд (2, 3, 4 или 5 мм). Двадцать четыре кролика были случайным образом отнесены к одной из этих групп, при этом два животных из группы 5 мм в конечном итоге были исключены из-за технических трудностей. Для каждого ежедневного сеанса УЗИ не проводилось в испытаниях, в которых ответ век достигал целевой амплитуды в пределах 10 мс после введения УЗИ.УЗИ проводилось в испытаниях, в которых реакция века не достигала целевой амплитуды.
Как показано на, животные в четырех группах приобрели устойчивые CR, и для каждой группы средняя амплитуда / величина ответов приближается к заданной цели. Для измерения амплитуды были включены все ответы, тогда как при измерении амплитуды были исключены все не-CR (те, у которых амплитуда <0,3 мм). Каждый из отдельных двухсторонних смешанных дисперсионных анализов (один для амплитуды и один для величины) показал значительную группу (амплитуда, F (3,198) = 7.43, p = 0,0039; величина, F = 11,18, p = 0,00013), тренировочный день (амплитуда, F (9,162) = 126,4, p <0,0001; величина, F = 101,21, p <0,0001 ) и взаимодействие группа × день (амплитуда, F (27,162) = 2,99, p <0,0001; величина, F = 3,66, p <0,0001). Эти анализы показывают, что животные учились, что между группами были различия в амплитуде и величине ответов, и что эти различия развивались в процессе обучения.Несмотря на эти различия в амплитуде и величине ответа, группы были сопоставимы по доле испытаний, в которых произошел полный ответ ( C ). Для этих данных двусторонний смешанный дисперсионный анализ показал значительный эффект дня тренировки ( F (9,162) = 123,8, p <0,0001), но не было никаких эффектов взаимодействия группы или группы × день тренировки ( p > 0,05).
Каждый из отдельных двухсторонних смешанных дисперсионных анализов (один для амплитуды и один для величины) показал значительную группу (амплитуда, F (3,198) = 7.43, p = 0,0039; величина, F = 11,18, p = 0,00013), тренировочный день (амплитуда, F (9,162) = 126,4, p <0,0001; величина, F = 101,21, p <0,0001 ) и взаимодействие группа × день (амплитуда, F (27,162) = 2,99, p <0,0001; величина, F = 3,66, p <0,0001). Эти анализы показывают, что животные учились, что между группами были различия в амплитуде и величине ответов, и что эти различия развивались в процессе обучения.Несмотря на эти различия в амплитуде и величине ответа, группы были сопоставимы по доле испытаний, в которых произошел полный ответ ( C ). Для этих данных двусторонний смешанный дисперсионный анализ показал значительный эффект дня тренировки ( F (9,162) = 123,8, p <0,0001), но не было никаких эффектов взаимодействия группы или группы × день тренировки ( p > 0,05).
Получение условных ответов век за 10 дней обучения для четырех групп, которым были назначены разные целевые амплитуды (2, 3, 4 и 5 мм). A , Амплитуда условных ответов век за 10 дней тренировки показывает получение разных амплитуд ответов для четырех групп. B , Аналогичные результаты были получены для измерения величины отклика, которая исключила ответы, которые не достигли критерия 0,3 мм, чтобы считаться CR. C , Поскольку вероятность ответа была устойчивой для всех четырех групп, результаты для амплитуды и величины были довольно схожими.В случаях, когда планки погрешностей (± SEM) не видны, ошибка меньше символа.
Есть два дополнительных индикатора, которые подтверждают гипотезу о том, что эта процедура ставит амплитуду ответа под экспериментальный контроль и более точно соответствует естественным условиям. Во-первых, должно быть уменьшение доли испытаний, в которых проводится УЗИ, с почти 1,0 до среднего уровня, что указывает на то, что амплитуда ответа иногда больше, а иногда меньше целевого. A показывает, что действительно доля испытаний, в которых выполнялась УЗИ, уменьшилась по сравнению с тренировкой, что указывает на увеличение количества испытаний, в которых целевая амплитуда была достигнута или превышена. Для этого показателя наблюдалось значительное влияние групп ( F (3,198) = 7,13, p = 0,0047) и тренировочного дня ( F (9,162) = 70,93, p <0,0001 ), но сравнение взаимодействий не было статистически надежным ( F (27,162) = 1.67, p = 0,0545). Во-вторых, должно происходить уменьшение средней ошибки, то есть среднего абсолютного значения разницы между амплитудой отклика и целевой амплитудой. Этот показатель отображается как функция количества тренировочных дней в B . На общее уменьшение средней ошибки указывает значительный эффект тренировочных дней ( F (9 162) = 102,17, p <0,0001). Поскольку начальное значение ошибки различается для четырех групп, неудивительно, что также наблюдался значительный групповой эффект ( F (3,198) = 14.
A показывает, что действительно доля испытаний, в которых выполнялась УЗИ, уменьшилась по сравнению с тренировкой, что указывает на увеличение количества испытаний, в которых целевая амплитуда была достигнута или превышена. Для этого показателя наблюдалось значительное влияние групп ( F (3,198) = 7,13, p = 0,0047) и тренировочного дня ( F (9,162) = 70,93, p <0,0001 ), но сравнение взаимодействий не было статистически надежным ( F (27,162) = 1.67, p = 0,0545). Во-вторых, должно происходить уменьшение средней ошибки, то есть среднего абсолютного значения разницы между амплитудой отклика и целевой амплитудой. Этот показатель отображается как функция количества тренировочных дней в B . На общее уменьшение средней ошибки указывает значительный эффект тренировочных дней ( F (9 162) = 102,17, p <0,0001). Поскольку начальное значение ошибки различается для четырех групп, неудивительно, что также наблюдался значительный групповой эффект ( F (3,198) = 14. 41, p <0,0001) и значительный эффект группы × тренировочные дни ( F (27,162) = 13,6, p <0,0001).
41, p <0,0001) и значительный эффект группы × тренировочные дни ( F (27,162) = 13,6, p <0,0001).
A , B , Уменьшение вероятности доставки в США ( A ) и средней ошибки ( B ) в зависимости от 10 дней обучения для четыре группы амплитуд (2, 3, 4 и 5 мм). Снижение вероятности УЗИ ( A ) указывает на увеличение доли ответов, которые соответствуют целевой амплитуде или превышают ее.Все четыре группы показали значительное снижение вероятности поражения в США, достигнув значений от 21 до 44%. B , Средняя ошибка уменьшилась до аналогичных значений во всех четырех группах.
В целом, эти анализы показывают, что (1) группы, разработанные с разницей в тренировках по амплитуде и величине ответа, (2) доля испытаний, в которых проводилось УЗИ, и средняя ошибка уменьшались за время обучения, и (3) там Нет различий в вероятности ответа для трех групп. Поскольку наиболее простой способ оценить свойства этого обучения – это оценить производительность в стабильном или близком к нему состоянии, далее мы сосредоточили анализ на данных за последние 2 дня обучения.
Поскольку наиболее простой способ оценить свойства этого обучения – это оценить производительность в стабильном или близком к нему состоянии, далее мы сосредоточили анализ на данных за последние 2 дня обучения.
Анализ асимптотической производительности
Для анализа стабильной производительности данные были объединены с 9-го и 10-го дней сбора данных для каждого субъекта. Этот анализ выявил четкие различия в амплитуде и величине CR между четырьмя группами ( A , B ).Отдельные односторонние дисперсионные анализы для амплитуды ( F (3,18) = 19,53, p <0,0001) и величины ( F = 37,25, p <0,0001) подтверждают надежность этих различий. Для измерения величины все шесть парных сравнений (критерий Тьюки, p <0,05) были надежными, а для амплитуды только сравнение 3 и 4 мм не было надежным. В отличие от этих различий, не было достоверных различий в измерении средней ошибки между четырьмя группами ( C ). В качестве финального теста на различия в амплитуде и величине ответов по четырем целевым амплитудным группам были рассчитаны нормализованные совокупные частотные распределения для каждой меры ответа ( G ). Тест Колмогорова – Смирнова ( p <0,005) показал, что все шесть парных сравнений для каждой группы были статистически надежными.
В качестве финального теста на различия в амплитуде и величине ответов по четырем целевым амплитудным группам были рассчитаны нормализованные совокупные частотные распределения для каждой меры ответа ( G ). Тест Колмогорова – Смирнова ( p <0,005) показал, что все шесть парных сравнений для каждой группы были статистически надежными.
Сравнения четырех амплитудных групп при стабильных показателях, взятых для каждого животного с 9-го и 10-го дней. A , Установившаяся амплитуда ответа различалась для четырех групп.Четыре пунктирные линии указывают целевую амплитуду для каждой группы. Все попарные сравнения (критерий Тьюки, p <0,05) были значимыми, за исключением сравнения между группами 3 и 4 мм. B , тот же формат, что и A для данных величины, которые представляют собой среднюю амплитуду всех CR без исключения не-CR. Все попарные сравнения были значимыми (критерий Тьюки, p <0,05). C , Стационарная средняя ошибка (абсолютное значение) существенно не различалась для четырех групп. D , Наблюдалась общая тенденция увеличения доли испытаний, в которых использовались УЗИ, с увеличением амплитуды цели. Единственное парное сравнение, которое было статистически надежным, было 2 против 5 мм. E , Все четыре группы показывают сопоставимые характеристики с точки зрения вероятности CR; между четырьмя группами не было значительных различий. F , Была общая тенденция к увеличению пиковой скорости CR по мере увеличения целевой амплитуды.Единственное достоверное (критерий Тьюки, p <0,05) сравнение было 2 против 5 мм. G , Суммарная доля ответов как функция амплитуды условного ответа (вверху) и величины (внизу) для четырех групп. Каждый CR для каждого животного помещали в бункер (ширина бункера 0,2 мм). Все шесть парных сравнений являются статистически надежными (критерий Колмогорова – Смирнова, p <0,005) для амплитуды, а также для измерения величины (кроме 4 против 5 мм, p <0.
C , Стационарная средняя ошибка (абсолютное значение) существенно не различалась для четырех групп. D , Наблюдалась общая тенденция увеличения доли испытаний, в которых использовались УЗИ, с увеличением амплитуды цели. Единственное парное сравнение, которое было статистически надежным, было 2 против 5 мм. E , Все четыре группы показывают сопоставимые характеристики с точки зрения вероятности CR; между четырьмя группами не было значительных различий. F , Была общая тенденция к увеличению пиковой скорости CR по мере увеличения целевой амплитуды.Единственное достоверное (критерий Тьюки, p <0,05) сравнение было 2 против 5 мм. G , Суммарная доля ответов как функция амплитуды условного ответа (вверху) и величины (внизу) для четырех групп. Каждый CR для каждого животного помещали в бункер (ширина бункера 0,2 мм). Все шесть парных сравнений являются статистически надежными (критерий Колмогорова – Смирнова, p <0,005) для амплитуды, а также для измерения величины (кроме 4 против 5 мм, p <0. 05).
05).
A – C в совокупности предполагают, что, хотя средняя ошибка амплитуды была сопоставима в четырех группах, источники ошибок были разными. Ответы для групп 2 и 3 мм имели тенденцию выходить за цель, тогда как ответы для группы 5 мм имели тенденцию выходить за пределы цели. Эту тенденцию можно вывести из сравнения вероятности в США, показанного в D , которое отражает долю ответов, которые не достигли целевого значения. Несмотря на четкую тенденцию к увеличению этого показателя с целевой амплитудой, различия между группами только приближались к значимости ( F (3,18) = 3.44, p = 0,08). Только группа 2 мм показала значительную тенденцию к превышению целевого значения, потому что отдельные тесты t для каждой группы выявили только достоверное отличие от среднего значения популяции в 50% (равные реакции недостижения и превышения) для группы 2 мм ( t ). (5) = 3,47, p <0,05). Несмотря на разницу в амплитуде и величине ответа, группы показали сопоставимые показатели ( F (3,18) = 1,799, p = 0. 37) с точки зрения вероятности CR ( E ). Наконец, F показывает, что существует также общая тенденция увеличения максимальной скорости КЛ с целевой амплитудой ( F (3,18) = 7,81, p = 0,003). Парные сравнения показали, что различия 2 по сравнению с 4 мм и 2 по сравнению с 5 мм были надежными (критерий Тьюки, p <0,05).
37) с точки зрения вероятности CR ( E ). Наконец, F показывает, что существует также общая тенденция увеличения максимальной скорости КЛ с целевой амплитудой ( F (3,18) = 7,81, p = 0,003). Парные сравнения показали, что различия 2 по сравнению с 4 мм и 2 по сравнению с 5 мм были надежными (критерий Тьюки, p <0,05).
Предыдущие исследования показали, что точное время CR является характерной чертой кондиционирования век (Smith, 1968; Smith et al., 1969; Милленсон и др., 1977; Маук и Руис, 1992). Время ожидания и время нарастания ответов систематически меняются в зависимости от межстимульного интервала, так что ответы имеют тенденцию достигать пика около времени доставки УЗИ. Мы исследовали различные меры времени ответа для четырех целевых амплитудных групп и не обнаружили достоверных различий во времени ответа ( A ). Ни время ожидания начала, ни время ожидания критерия, время ожидания максимальной скорости, ни время ожидания ответа пика не показали статистически достоверных различий между группами. Когда ответы были разделены в зависимости от того, превышают ли они цель (испытания в США) или ниже цели (испытания в США), наблюдались значительные различия во времени ответов во всех четырех группах ( B ). Отдельные двухсторонние смешанные ANOVA для задержки начала и задержки до максимальной скорости показали только значимое влияние типа ответа (не в США по сравнению с США; начало, F (1,18) = 100,6, p < 0,0001; пиковая скорость, F = 49.04, p <0,0001). Этот анализ показывает, что в целом ответы, которые соответствовали целевой амплитуде или превышали ее, имели более короткие задержки, чем ответы, которые не соответствовали целевой амплитуде.
Когда ответы были разделены в зависимости от того, превышают ли они цель (испытания в США) или ниже цели (испытания в США), наблюдались значительные различия во времени ответов во всех четырех группах ( B ). Отдельные двухсторонние смешанные ANOVA для задержки начала и задержки до максимальной скорости показали только значимое влияние типа ответа (не в США по сравнению с США; начало, F (1,18) = 100,6, p < 0,0001; пиковая скорость, F = 49.04, p <0,0001). Этот анализ показывает, что в целом ответы, которые соответствовали целевой амплитуде или превышали ее, имели более короткие задержки, чем ответы, которые не соответствовали целевой амплитуде.
Меры времени отклика по сравнению с четырьмя группами амплитуды. A , Четыре графика показывают задержку до начала, задержку до критерия, задержку до пиковой скорости и задержку до пиковой амплитуды (для испытаний за пределами США). Статистически достоверной разницы между группами по этим показателям ответа не было. B , Разница во времени испытаний в США и за пределами США. Раздельный анализ исследований в УЗИ (амплитуда ниже целевого) и неамериканских исследований (амплитуда на уровне или выше) выявил общую тенденцию к более быстрым и коротким задержкам ответов в неамериканских исследованиях. Все восемь сравнений (4 группы × 2 меры ответа) были статистически надежными (парный тест t , p <0,01).
B , Разница во времени испытаний в США и за пределами США. Раздельный анализ исследований в УЗИ (амплитуда ниже целевого) и неамериканских исследований (амплитуда на уровне или выше) выявил общую тенденцию к более быстрым и коротким задержкам ответов в неамериканских исследованиях. Все восемь сравнений (4 группы × 2 меры ответа) были статистически надежными (парный тест t , p <0,01).
Примеры необработанных ответов от одного испытуемого из каждой амплитудной группы (день 10 в каждом случае) показаны в.Для каждого субъекта ответы, которые соответствовали целевой амплитуде или превышали ее, показаны в левом столбце, а те, которые не соответствовали целевой амплитуде (и, следовательно, были доставлены УЗИ), показаны в правом столбце. Эти данные показывают общую тенденцию реакции к перерегулированию для меньшей амплитуды цели и к недолету для цели 5 мм. Вместе эти анализы демонстрируют, что процедура тренировки целевой амплитуды может использоваться для получения разумного экспериментального контроля над амплитудой ответа при кондиционировании век, что средняя ошибка амплитуды сопоставима для целей от 2 до 5 мм, и что эти различия возникают на сопоставимых уровнях CR. и сопоставимые статистические данные о сроках проведения CR.
и сопоставимые статистические данные о сроках проведения CR.
Пример необработанных ответов от четырех животных, по одному от каждой группы амплитуд, взятых с 10-го дня обучения. Каждая развертка представляет собой отдельную реакцию века (первая проба впереди), причем черная область каждой развертки указывает продолжительность CS. Ответы были разделены на испытания, проводимые за пределами США (левый столбец) и испытания в США (правый столбец). Последовательность испытаний каждого животного в США и за пределами США очевидна по интервалу между испытаниями. Обратите внимание на общую тенденцию к большему количеству испытаний без УЗИ с малыми амплитудами цели и к большему количеству испытаний УЗИ с большими амплитудами цели.Компонент рефлекторного ответа каждого испытания виден в светло-серой области испытаний «With-US».
Амплитуда отклика после повторного обнаружения новой цели
Практическая и концептуальная важность тренировки амплитуды целевой будет уменьшена, если амплитуды отклика цели не могут быть получены и повторно получены в соответствии с планом эксперимента. Поэтому мы тестировали каждое животное, используя новую целевую амплитуду, отличную от начальной. После 10 дней обучения до начальной целевой амплитуды каждый субъект был обучен в течение 10 дополнительных дней с использованием новой целевой амплитуды.Новые целевые амплитуды для групп 2, 3, 4 и 5 мм, соответственно, составляли 4, 5, 2 и 3 мм. показывает сравнение амплитуд ответа для этих четырех групп для начальной и новой целевой амплитуд. Как и выше, эти данные включали объединенное среднее значение 9 и 10 дней для каждого субъекта. Мы начали с того, что спросили, есть ли различия между получением и повторным обнаружением для каждой из четырех целевых амплитуд (левый график). Это сравнение между субъектами сравнивает каждую цель, когда она использовалась при захвате, по сравнению с повторным захватом (т.е., получение от группы 2 мм и повторное обнаружение на цели 2 мм, первоначально группы 4 мм). Двусторонний дисперсионный анализ выявил только основной эффект целевой амплитуды ( F (3,36) = 37,06, p <0,0001), эффекты захвата по сравнению с повторным захватом и взаимодействия не были значительными, и не было надежных различия в амплитудах, полученных во время захвата, по сравнению с повторным обнаружением новой цели.
Поэтому мы тестировали каждое животное, используя новую целевую амплитуду, отличную от начальной. После 10 дней обучения до начальной целевой амплитуды каждый субъект был обучен в течение 10 дополнительных дней с использованием новой целевой амплитуды.Новые целевые амплитуды для групп 2, 3, 4 и 5 мм, соответственно, составляли 4, 5, 2 и 3 мм. показывает сравнение амплитуд ответа для этих четырех групп для начальной и новой целевой амплитуд. Как и выше, эти данные включали объединенное среднее значение 9 и 10 дней для каждого субъекта. Мы начали с того, что спросили, есть ли различия между получением и повторным обнаружением для каждой из четырех целевых амплитуд (левый график). Это сравнение между субъектами сравнивает каждую цель, когда она использовалась при захвате, по сравнению с повторным захватом (т.е., получение от группы 2 мм и повторное обнаружение на цели 2 мм, первоначально группы 4 мм). Двусторонний дисперсионный анализ выявил только основной эффект целевой амплитуды ( F (3,36) = 37,06, p <0,0001), эффекты захвата по сравнению с повторным захватом и взаимодействия не были значительными, и не было надежных различия в амплитудах, полученных во время захвата, по сравнению с повторным обнаружением новой цели. Правая часть показывает те же данные, построенные заново, чтобы выделить сравнения амплитуд, полученных для сбора и повторного сбора данных в одних и тех же группах.Четыре отдельных парных теста t ( p <0,01) показали, что для каждой группы амплитуда ответов, полученных при обнаружении одной цели, отличалась от тех, которые были получены при повторном обнаружении на новую цель. Эти анализы показывают, что как получение первой целевой амплитуды, так и повторное обнаружение новой целевой амплитуды вызвали значительную разницу в амплитуде ответа для каждой целевой амплитудной группы и что не было никаких различий в этих тенденциях между захватом и повторным захватом.
Правая часть показывает те же данные, построенные заново, чтобы выделить сравнения амплитуд, полученных для сбора и повторного сбора данных в одних и тех же группах.Четыре отдельных парных теста t ( p <0,01) показали, что для каждой группы амплитуда ответов, полученных при обнаружении одной цели, отличалась от тех, которые были получены при повторном обнаружении на новую цель. Эти анализы показывают, что как получение первой целевой амплитуды, так и повторное обнаружение новой целевой амплитуды вызвали значительную разницу в амплитуде ответа для каждой целевой амплитудной группы и что не было никаких различий в этих тенденциях между захватом и повторным захватом.
Сравнение устойчивых амплитуд ответа после первоначального сбора данных (темно-серый; те же данные, что и A ) и у тех же животных после 10 дней повторного обнаружения с новой целевой амплитудой. Начальная амплитуда цели и амплитуда цели повторного обнаружения были, соответственно, для четырех групп (от 2 до 4 мм, от 3 до 5 мм, от 4 до 2 мм и от 5 до 3 мм). Как и в случае, данные получены из среднего значения для каждого животного за 9-й и 10-й дни захвата (и повторного захвата). Слева: сравнение идентичных целевых амплитуд для захвата и повторного захвата.Статистически достоверных различий для каждой целевой амплитуды между данными сбора и повторного обнаружения не было ( t тестов). Целевая амплитуда обозначена пунктирными линиями. Справа: сравнение данных приобретения и повторного приобретения для одних и тех же групп животных. Различные целевые амплитуды захвата и повторного захвата для каждой группы снова обозначены пунктирными линиями. Парные тесты t ( p <0,01) показали статистически достоверные различия между амплитудами сбора и повторного обнаружения для каждой группы.
Как и в случае, данные получены из среднего значения для каждого животного за 9-й и 10-й дни захвата (и повторного захвата). Слева: сравнение идентичных целевых амплитуд для захвата и повторного захвата.Статистически достоверных различий для каждой целевой амплитуды между данными сбора и повторного обнаружения не было ( t тестов). Целевая амплитуда обозначена пунктирными линиями. Справа: сравнение данных приобретения и повторного приобретения для одних и тех же групп животных. Различные целевые амплитуды захвата и повторного захвата для каждой группы снова обозначены пунктирными линиями. Парные тесты t ( p <0,01) показали статистически достоверные различия между амплитудами сбора и повторного обнаружения для каждой группы.
Стимуляция мшистых волокон в качестве CS
Значительные доказательства указывают на то, что приобретение и экспрессия CR во время формирования павлового (отложенного) века опосредуется пластичностью мозжечка. Поскольку обусловленность целевой амплитуды, строго говоря, не является павловской, мы провели два дополнительных эксперимента, чтобы проверить возможность того, что обучение в этих условиях опосредуется нецеребеллярными механизмами. Способность заменять стимуляцию мшистых волокон тонусом CS – одно из наиболее убедительных доказательств, подтверждающих, что мозжечковые механизмы являются основой кондиционирования век.Таким образом, мы сравнили эффекты тренировки с целевой амплитудой 3 мм, используя тон CS в одной группе и стимуляцию мшистых волокон в качестве CS во второй группе. В остальном процедуры были идентичны процедурам, использованным выше для группы 3 мм. показывает результаты за 10 дней тренировок для этих двух групп. A – D сравнивает эти две группы за 10 дней обучения, а E показывает сравнения в установившемся состоянии, как в предыдущем эксперименте. Отдельные двухфакторные смешанные ANOVA для четырех мер реакции показали только основные эффекты тренировочных дней (амплитуда, F (9,54) = 30.54; ошибка, F = 17,71; Вероятность CR, F = 29,09; Вероятность США – 16,62; p <0,0001 во всех случаях). Ни один из основных эффектов группы или эффектов взаимодействия не был значимым, равно как и сравнение тона и мшистого волокна ( E ) для восьми показателей реакции, которые были проанализированы для данных устойчивого состояния (дни 9 и 10, t тесты, p > 0,05).
Способность заменять стимуляцию мшистых волокон тонусом CS – одно из наиболее убедительных доказательств, подтверждающих, что мозжечковые механизмы являются основой кондиционирования век.Таким образом, мы сравнили эффекты тренировки с целевой амплитудой 3 мм, используя тон CS в одной группе и стимуляцию мшистых волокон в качестве CS во второй группе. В остальном процедуры были идентичны процедурам, использованным выше для группы 3 мм. показывает результаты за 10 дней тренировок для этих двух групп. A – D сравнивает эти две группы за 10 дней обучения, а E показывает сравнения в установившемся состоянии, как в предыдущем эксперименте. Отдельные двухфакторные смешанные ANOVA для четырех мер реакции показали только основные эффекты тренировочных дней (амплитуда, F (9,54) = 30.54; ошибка, F = 17,71; Вероятность CR, F = 29,09; Вероятность США – 16,62; p <0,0001 во всех случаях). Ни один из основных эффектов группы или эффектов взаимодействия не был значимым, равно как и сравнение тона и мшистого волокна ( E ) для восьми показателей реакции, которые были проанализированы для данных устойчивого состояния (дни 9 и 10, t тесты, p > 0,05). Эти данные исключают факторы, расположенные выше мозжечка, как способствующие приобретению амплитуды ответа, и подтверждают гипотезу о том, что кондиционирование век к целевой амплитуде опосредуется мозжечком.
Эти данные исключают факторы, расположенные выше мозжечка, как способствующие приобретению амплитуды ответа, и подтверждают гипотезу о том, что кондиционирование век к целевой амплитуде опосредуется мозжечком.
Получение целевой амплитуды (3 мм) для животных, обученных с помощью тонального CS и стимуляции мшистыми волокнами в качестве CS. A , Амплитуда CR за 10 дней обучения. B , Средняя ошибка (абсолютное значение) за 10 дней обучения. MF, Мшистое волокно. C , D , вероятность CR ( C ) и вероятность США ( D ) за 10 дней обучения. E , Сравнение между группами CS тональной стимуляции и CS со стимуляцией мшистыми волокнами для различных мер реакции, взятых с 9-го и 10-го дней тренировки.Ни одно из шести сравнений групп не было статистически надежным (тест t , p > 0,05).
Обратимая инактивация мозжечка
В качестве дополнительной проверки гипотезы о том, что мозжечок необходим для кондиционирования до заданной амплитуды, экспрессия CR была протестирована на четырех животных, обученных тону, и четырех животных, обученных мшистым волокнам, до и после инактивации. мозжечкового выхода через инфузию мусцимола, агониста ГАМК, в переднее межположительное ядро ().Субъекты в обеих группах тестировались в отдельные дни с инфузиями носителя или инфузиями мусцимола (порядок уравновешивался для каждой группы). График средних показателей по группе (в центре) показывает, что для обеих групп не было достоверных изменений после инфузии носителя, тогда как инфузии мусцимола отменяли экспрессию CR. Необработанные данные для одного субъекта стимуляции мшистых волокон для инфузий носителя (слева) и мусцимола (справа) показаны вверху. Пример канюли (слева) и стимулирующих электродов от того же пациента показаны внизу.
Необходимость мозжечка для выражения приобретенных ответов на заданную амплитуду 3 мм. Две группы, одна из которых использовала тональный CS, а другая, в которой стимуляция волокон использовалась в качестве CS, были протестированы в отдельные дни либо с инфузией носителя, либо с инфузией мусцимола, агониста ГАМК. На двух графиках показаны средние показатели по группе отсутствия эффекта от инфузии носителя (слева) и отмены CR с помощью инфузии мусцимола (справа). Время настаивания в каждом случае указано черной стрелкой.Примеры исходных данных от одного животного показаны выше: следы слева – это инфузия носителя, а следы справа – инфузия мусцимола. В каждом случае ответы до инфузии находятся впереди (первое испытание впереди), а реакции после инфузии – сзади. На двух нижних фотографиях показано размещение инфузионной канюли (слева) и стимулирующих электродов из мшистых волокон (справа) для животного, исходные данные которого показаны выше. Этому животному были имплантированы два электрода из покрытых мхом волокон; только один был использован для этого эксперимента.MF, Мшистые волокна.
Две группы, одна из которых использовала тональный CS, а другая, в которой стимуляция волокон использовалась в качестве CS, были протестированы в отдельные дни либо с инфузией носителя, либо с инфузией мусцимола, агониста ГАМК. На двух графиках показаны средние показатели по группе отсутствия эффекта от инфузии носителя (слева) и отмены CR с помощью инфузии мусцимола (справа). Время настаивания в каждом случае указано черной стрелкой.Примеры исходных данных от одного животного показаны выше: следы слева – это инфузия носителя, а следы справа – инфузия мусцимола. В каждом случае ответы до инфузии находятся впереди (первое испытание впереди), а реакции после инфузии – сзади. На двух нижних фотографиях показано размещение инфузионной канюли (слева) и стимулирующих электродов из мшистых волокон (справа) для животного, исходные данные которого показаны выше. Этому животному были имплантированы два электрода из покрытых мхом волокон; только один был использован для этого эксперимента.MF, Мшистые волокна.
В совокупности исследования стимуляции мшистых волокон и инактивации глубоких ядер показывают (1), что мозжечок необходим для экспрессии CR, продуцируемых кондиционированием века до целевой амплитуды (результаты инфузии), и (2) что механизмы, ответственные за приобретение и экспрессия целевой амплитуды ответа не находятся в пути CR выше мозжечка (результаты стимуляции мшистыми волокнами).
Обсуждение
Мы адаптировали традиционные процедуры кондиционирования павловских век, чтобы ввод ошибки, определяемый США, зависел от амплитуды CR.Пропустив УЗИ в исследованиях, в которых CR соответствует или превышает заданную целевую амплитуду (и представив УЗИ в противном случае), мы показали, что результирующие условные реакции век имеют тенденцию к целевой амплитуде. Это верно как для получения исходной целевой амплитуды, так и для повторного обнаружения с использованием другой цели. Несмотря на то, что наблюдалась тенденция к перерегулированию при малых амплитудах цели и к недолету с наибольшей целью, средняя абсолютная ошибка не различалась для групп, обученных для целевых амплитуд от 2 до 5 мм. Эти данные показывают, что, адаптируя процедуры кондиционирования век к естественным условиям обучения мозжечка, амплитуду CR можно поставить под экспериментальный контроль. Способность устранять эти CR с помощью инактивации мозжечка и способность заменять стимуляцию мшистых волокон тональным CS подтверждают гипотезу о том, что, как и в случае кондиционирования века в форме павлова, кондиционирование века до целевой амплитуды опосредуется мозжечком. Эти результаты формируют основу для систематических и контролируемых исследований того, как мозжечок обучается, сохраняет и выражает надлежащую амплитуду откликов, которые он адаптирует.
Эти данные показывают, что, адаптируя процедуры кондиционирования век к естественным условиям обучения мозжечка, амплитуду CR можно поставить под экспериментальный контроль. Способность устранять эти CR с помощью инактивации мозжечка и способность заменять стимуляцию мшистых волокон тональным CS подтверждают гипотезу о том, что, как и в случае кондиционирования века в форме павлова, кондиционирование века до целевой амплитуды опосредуется мозжечком. Эти результаты формируют основу для систематических и контролируемых исследований того, как мозжечок обучается, сохраняет и выражает надлежащую амплитуду откликов, которые он адаптирует.
Значение для кондиционирования век и мозжечка
Необходимость мозжечка для кондиционирования века с задержкой павлова подтверждается большим количеством данных. Настоящие результаты показывают, что непредвиденные обстоятельства Павлова, при которых доставка УЗИ не зависит от CR, не являются особенностью, которая делает мозжечок необходимым. Поскольку, строго говоря, процедуры тренировки до целевой амплитуды включают в себя оперантные непредвиденные обстоятельства, необходимость мозжечка в кондиционировании век связана с природой стимулов и системой реакции, а не с природой непредвиденных обстоятельств, которые управляют представлением усиливающий стимул.
Настоящие данные также обращаются к распространенному заблуждению о кондиционировании век, а именно, что это рефлекторное кондиционирование или кондиционирование моргания глаз. Павлов выступил с идеей, что классическое кондиционирование включает в себя пути, активируемые CS, обретающие способность активировать путь, который опосредует рефлекторный ответ (Павлов, 1927). Нервные пути, которые, как известно, опосредуют экспрессию CR и рефлекторные мигания, вызываемые США, не согласуются с гипотезой Павлова. Рефлекторные мигания, вызываемые первичными афферентами, проходящими через верхний глазничный нерв, опосредуются двумя путями (Pellegrini et al., 1995). Первичные афференты проецируются как на каудально-межполярную пограничную область спинномозгового ядра тройничного нерва, так и на дорсальный рог, сегмент С1 спинного мозга. Обе эти области проецируются в область orbicularis oculi лицевого ядра. Эксперименты с поражением показали, что спинальный тройничный путь опосредует начальный компонент R1 рефлекторных морганий, а путь спинного мозга опосредует более длинный латентный компонент R2 (Pelligrini et al. , 1995). Путь CR, выступающий из переднего межположительного ядра через красное ядро, по-видимому, соединяется с этим путем в двигательных или премоторных нейронах лицевого ядра.Следовательно, рефлекторные мигания и CRs опосредуются совершенно разными нервными путями и, как можно ожидать, будут проявлять совершенно разные свойства, как было четко установлено в исследованиях на людях (Schade Powers et al., 2010). Поэтому условные реакции век более правильно охарактеризовать как «реакции век», чем «моргания глаз», и настоящие данные демонстрируют, что амплитуда этих ответов век, как и большинство других реакций скелетных мышц, может быть изучена и опосредована обучением мозжечка.
, 1995). Путь CR, выступающий из переднего межположительного ядра через красное ядро, по-видимому, соединяется с этим путем в двигательных или премоторных нейронах лицевого ядра.Следовательно, рефлекторные мигания и CRs опосредуются совершенно разными нервными путями и, как можно ожидать, будут проявлять совершенно разные свойства, как было четко установлено в исследованиях на людях (Schade Powers et al., 2010). Поэтому условные реакции век более правильно охарактеризовать как «реакции век», чем «моргания глаз», и настоящие данные демонстрируют, что амплитуда этих ответов век, как и большинство других реакций скелетных мышц, может быть изучена и опосредована обучением мозжечка.
Значение для обучения мозжечка усилению или амплитуде
Вопрос о том, как и где мозжечок обучается и сохраняет усиление или амплитуду ответа, взаимосвязан с вопросом о месте (ах) пластичности, ответственных за обучение мозжечка. На этот вопрос лучше всего ответить с помощью исследований, включающих анализ адаптации VOR (Raymond et al. , 1996; Boyden et al., 2004). Первоначальные усилия в этой области вызвали серьезные дискуссии о том, происходит ли пластичность в коре мозжечка или в вестибулярном ядре, чьи связи и схема параллельны глубоким ядрам мозжечка (Lisberger and Sejnowski, 1992, 1993; Ito, 1993).Более поздние теории предполагают участие пластичности как в коре мозжечка, так и в глубоком ядре (Perrett et al., 1993; Raymond et al., 1996; Mauk and Donegan, 1997). Когда задействовано более одного сайта пластичности, становится важным вопрос об относительном вкладе этих двух сайтов. Относительный вклад коры мозжечка и механизмов пластичности глубокого ядра мозжечка в усиление или амплитуду ответа остается нерешенным, и есть указания на то, что относительный вклад может варьироваться сложным образом в зависимости от обстоятельств (Mauk, 1997).Кондиционирование век до целевой амплитуды обеспечивает способ решения этих вопросов с помощью относительно простой и экспериментально управляемой системы, которая поддается параметрическому поведенческому анализу, обратимым повреждениям, in vivo записи и даже крупномасштабному компьютерному моделированию.
, 1996; Boyden et al., 2004). Первоначальные усилия в этой области вызвали серьезные дискуссии о том, происходит ли пластичность в коре мозжечка или в вестибулярном ядре, чьи связи и схема параллельны глубоким ядрам мозжечка (Lisberger and Sejnowski, 1992, 1993; Ito, 1993).Более поздние теории предполагают участие пластичности как в коре мозжечка, так и в глубоком ядре (Perrett et al., 1993; Raymond et al., 1996; Mauk and Donegan, 1997). Когда задействовано более одного сайта пластичности, становится важным вопрос об относительном вкладе этих двух сайтов. Относительный вклад коры мозжечка и механизмов пластичности глубокого ядра мозжечка в усиление или амплитуду ответа остается нерешенным, и есть указания на то, что относительный вклад может варьироваться сложным образом в зависимости от обстоятельств (Mauk, 1997).Кондиционирование век до целевой амплитуды обеспечивает способ решения этих вопросов с помощью относительно простой и экспериментально управляемой системы, которая поддается параметрическому поведенческому анализу, обратимым повреждениям, in vivo записи и даже крупномасштабному компьютерному моделированию.
Добавляя, сколько и когда
Abstract
Концептуальные и практические преимущества кондиционирования павловых век облегчают анализ вычислений и обучения мозжечка.Тем не менее, процедуры кондиционирования век во многом нереальны. Сигнал ошибки для оливоцеребеллярной системы не уменьшается, поскольку обучение адаптирует амплитуду или усиление ответа. Это по своей сути ограничивает полезность кондиционирования век для исследований, изучающих, как механизмы обучения мозжечка приобретают и сохраняют амплитуду адаптивного ответа. Мы сообщаем о разработке и описании процедуры обучения, в которой амплитуда условной реакции ставится под экспериментальный контроль с непредвиденными обстоятельствами, которые более близки к естественным условиям.В этой процедуре доставка безусловного стимула (US) зависит от амплитуды условного ответа: US доставляется для ответов, которые не достигают заданной целевой амплитуды, и опускается для ответов, которые соответствуют или превышают цель. Мы обнаружили, что кролики, обученные либо тональной стимуляции, либо стимуляции мшистыми волокнами в качестве условного стимула, усваивают ответы, которые достигают целевых амплитуд в диапазоне от 2 до 5 мм. Инактивация межположительного ядра с помощью инфузий мусцимола устраняет эти условные реакции, указывая на то, что участие мозжечка в кондиционировании век не связано явно с использованием процедур Павлова.Вместе с предыдущими исследованиями эти данные предполагают, что амплитуда ответа изучается и кодируется в мозжечке во время кондиционирования век. Таким образом, эти результаты обеспечивают основу для систематических и контролируемых исследований мозжечковых механизмов, которые обучаются и кодируют правильную амплитуду адаптивных движений.
Мы обнаружили, что кролики, обученные либо тональной стимуляции, либо стимуляции мшистыми волокнами в качестве условного стимула, усваивают ответы, которые достигают целевых амплитуд в диапазоне от 2 до 5 мм. Инактивация межположительного ядра с помощью инфузий мусцимола устраняет эти условные реакции, указывая на то, что участие мозжечка в кондиционировании век не связано явно с использованием процедур Павлова.Вместе с предыдущими исследованиями эти данные предполагают, что амплитуда ответа изучается и кодируется в мозжечке во время кондиционирования век. Таким образом, эти результаты обеспечивают основу для систематических и контролируемых исследований мозжечковых механизмов, которые обучаются и кодируют правильную амплитуду адаптивных движений.
Введение
Павловское кондиционирование века облегчило анализ обучения и вычислений мозжечка (McCormick and Thompson, 1984; Perrett et al., 1993; Маук и Донеган, 1997; Медина и Маук, 1999; Hansel et al., 2001; Medina et al., 2001, 2002), в основном из-за тесного соответствия между тренировочными стимулами и входными сигналами мозжечка, а также между выходной мощностью мозжечка и поведенческими реакциями (McCormick and Thompson, 1984; Mauk et al. , 1986; Steinmetz et al. , 1986; Mauk, Donegan, 1997; Hesslow et al., 1999). Идея Павлова (1927) о том, что условное кондиционирование – это «рефлексивное обучение», при котором условный стимул (КС) приобретает способность активировать рефлекторный путь, запускаемый подкрепляющим стимулом, является неправильным представлением, обычно применяемым к кондиционированию век.В самом деле, неправильное название «кондиционирование моргания», по-видимому, происходит из этого заблуждения. Данные, противоречащие этой идее, включают известные схемы кондиционирования век, которые ясно показывают, что путь CS не связан с рефлекторным путем через обучение (Mauk and Donegan, 1997; Thompson et al., 1998), из доказательств того, что условные реакции век не являются морганиями (Schade Powers et al., 2010), а из простого поведенческого наблюдения, что во время приобретения амплитуда условной реакции не является однозначной.С тренировкой амплитуда условной реакции монотонно растет от едва заметного до максимального закрытия век.
, 1986; Steinmetz et al. , 1986; Mauk, Donegan, 1997; Hesslow et al., 1999). Идея Павлова (1927) о том, что условное кондиционирование – это «рефлексивное обучение», при котором условный стимул (КС) приобретает способность активировать рефлекторный путь, запускаемый подкрепляющим стимулом, является неправильным представлением, обычно применяемым к кондиционированию век.В самом деле, неправильное название «кондиционирование моргания», по-видимому, происходит из этого заблуждения. Данные, противоречащие этой идее, включают известные схемы кондиционирования век, которые ясно показывают, что путь CS не связан с рефлекторным путем через обучение (Mauk and Donegan, 1997; Thompson et al., 1998), из доказательств того, что условные реакции век не являются морганиями (Schade Powers et al., 2010), а из простого поведенческого наблюдения, что во время приобретения амплитуда условной реакции не является однозначной.С тренировкой амплитуда условной реакции монотонно растет от едва заметного до максимального закрытия век.
Тем не менее, стандартные тренировочные процедуры кондиционирования век не позволяют экспериментально контролировать амплитуду ответа. Для более естественных реакций, таких как адаптация вестибулоокулярного рефлекса (VOR), вход ошибки в оливо-церебеллярную систему (т. Е. Входной сигнал восходящего волокна в мозжечок) уменьшается по мере того, как обучение адаптирует усиление или амплитуду ответа (Gilbert and Thach, 1977). ; Раймонд и Лисбергер, 1997, 1998).Поскольку вероятность сигнала ошибки в оливоцеребеллярной системе [через предъявление безусловного стимула (УЗ)] не уменьшается по мере роста амплитуды условного ответа, процедуры Павлова по своей природе способствуют условным ответам (КО) с максимальной амплитудой (5–6 мм для кроликов). ). Даже при воздушной затяжке, такой как УЗИ, CR (расширение мигательной перепонки) не может полностью устранить активацию УЗ-лазейных волокон, потому что внешние веки обычно ограничены, а сама мигательная перепонка весьма чувствительна.По этим причинам кондиционирование век не было полезным для анализа мозжечковых механизмов, участвующих в обучении и кодировании амплитуды ответа (то есть усиления ответа).
Мы сообщаем о разработке и описании процедуры тренировки век, которая более точно имитирует естественные требования к обучению мозжечка и ставит амплитуду ответа под экспериментальный контроль. В этой процедуре УЗИ зависит от амплитуды условного ответа, так же как ошибочные воздействия на мозжечок зависят от усиления ответа.Когда амплитуда ответа соответствует заранее заданной целевой амплитуде или превышает ее, УЗИ не выполняется (см.). Мы демонстрируем, что амплитуда ответа приближается к целевым амплитудам в диапазоне от 2 до 5 мм, со значительными групповыми различиями при анализе после 10 тренировок. Участие мозжечка в этой обучающей задаче подтверждается двумя наблюдениями: (1) амплитуда ответа может быть определена, когда стимуляция мшистых волокон заменяет тон, и (2) инактивация мозжечкового выхода путем вливания мусцимола в межположительное ядро устраняет экспрессию CR, что позволяет предположить что мозжечковое обучение отвечает не только за то, нужно ли и когда реагировать, но и за то, насколько сильно реагировать. Эти данные показывают, что эта новая тренировочная процедура представляет собой управляемую экспериментальную парадигму для изучения мозжечковых механизмов, которые обучаются и кодируют амплитуду ответа.
Эти данные показывают, что эта новая тренировочная процедура представляет собой управляемую экспериментальную парадигму для изучения мозжечковых механизмов, которые обучаются и кодируют амплитуду ответа.
Схематическое изображение тренировочных процедур, используемых для тренировки реакции век на заданную амплитуду. Каждое тренировочное испытание включало предъявление тонального сигнала CS и запланированную доставку ультразвуковой стимуляции глаза с межстимульным интервалом 500 мс. Фактическая доставка УЗИ зависела от амплитуды ответа путем мониторинга амплитуды ответа в реальном времени во время CS.За десять миллисекунд до запланированной доставки США (время принятия решения в США) обучающее программное обеспечение было запрограммировано таким образом, чтобы определять, будут ли доставлены УЗИ: для ответов, которые соответствовали или превышали заданную амплитуду, доставлялись УЗИ; для ответов, которые не достигли целевой амплитуды, доставлялся УЗИ.
Результаты
Получение целевой амплитуды отклика
В естественных условиях, таких как адаптация VOR, преследования или саккад, вызванное ошибкой возбуждающее воздействие на вход восходящего волокна в мозжечок уменьшается по мере того, как усиление отклика приближается к правильному значению (Гилберт и Тач, 1977; Раймонд и Лисбергер, 1997, 1998). Наша процедура обучения предназначена для более точного приближения к этим естественным условиям обучения путем связывания УЗ-презентации с амплитудой ответа относительно целевой амплитуды. Для этого мы тренировали четыре группы животных в течение 10 дней, каждая группа подвергалась одной из четырех целевых амплитуд (2, 3, 4 или 5 мм). Двадцать четыре кролика были случайным образом отнесены к одной из этих групп, при этом два животных из группы 5 мм в конечном итоге были исключены из-за технических трудностей. Для каждого ежедневного сеанса УЗИ не проводилось в испытаниях, в которых ответ век достигал целевой амплитуды в пределах 10 мс после введения УЗИ.УЗИ проводилось в испытаниях, в которых реакция века не достигала целевой амплитуды.
Наша процедура обучения предназначена для более точного приближения к этим естественным условиям обучения путем связывания УЗ-презентации с амплитудой ответа относительно целевой амплитуды. Для этого мы тренировали четыре группы животных в течение 10 дней, каждая группа подвергалась одной из четырех целевых амплитуд (2, 3, 4 или 5 мм). Двадцать четыре кролика были случайным образом отнесены к одной из этих групп, при этом два животных из группы 5 мм в конечном итоге были исключены из-за технических трудностей. Для каждого ежедневного сеанса УЗИ не проводилось в испытаниях, в которых ответ век достигал целевой амплитуды в пределах 10 мс после введения УЗИ.УЗИ проводилось в испытаниях, в которых реакция века не достигала целевой амплитуды.
Как показано на, животные в четырех группах приобрели устойчивые CR, и для каждой группы средняя амплитуда / величина ответов приближается к заданной цели. Для измерения амплитуды были включены все ответы, тогда как при измерении амплитуды были исключены все не-CR (те, у которых амплитуда <0,3 мм). Каждый из отдельных двухсторонних смешанных дисперсионных анализов (один для амплитуды и один для величины) показал значительную группу (амплитуда, F (3,198) = 7.43, p = 0,0039; величина, F = 11,18, p = 0,00013), тренировочный день (амплитуда, F (9,162) = 126,4, p <0,0001; величина, F = 101,21, p <0,0001 ) и взаимодействие группа × день (амплитуда, F (27,162) = 2,99, p <0,0001; величина, F = 3,66, p <0,0001). Эти анализы показывают, что животные учились, что между группами были различия в амплитуде и величине ответов, и что эти различия развивались в процессе обучения.Несмотря на эти различия в амплитуде и величине ответа, группы были сопоставимы по доле испытаний, в которых произошел полный ответ ( C ). Для этих данных двусторонний смешанный дисперсионный анализ показал значительный эффект дня тренировки ( F (9,162) = 123,8, p <0,0001), но не было никаких эффектов взаимодействия группы или группы × день тренировки ( p > 0,05).
Каждый из отдельных двухсторонних смешанных дисперсионных анализов (один для амплитуды и один для величины) показал значительную группу (амплитуда, F (3,198) = 7.43, p = 0,0039; величина, F = 11,18, p = 0,00013), тренировочный день (амплитуда, F (9,162) = 126,4, p <0,0001; величина, F = 101,21, p <0,0001 ) и взаимодействие группа × день (амплитуда, F (27,162) = 2,99, p <0,0001; величина, F = 3,66, p <0,0001). Эти анализы показывают, что животные учились, что между группами были различия в амплитуде и величине ответов, и что эти различия развивались в процессе обучения.Несмотря на эти различия в амплитуде и величине ответа, группы были сопоставимы по доле испытаний, в которых произошел полный ответ ( C ). Для этих данных двусторонний смешанный дисперсионный анализ показал значительный эффект дня тренировки ( F (9,162) = 123,8, p <0,0001), но не было никаких эффектов взаимодействия группы или группы × день тренировки ( p > 0,05).
Получение условных ответов век за 10 дней обучения для четырех групп, которым были назначены разные целевые амплитуды (2, 3, 4 и 5 мм). A , Амплитуда условных ответов век за 10 дней тренировки показывает получение разных амплитуд ответов для четырех групп. B , Аналогичные результаты были получены для измерения величины отклика, которая исключила ответы, которые не достигли критерия 0,3 мм, чтобы считаться CR. C , Поскольку вероятность ответа была устойчивой для всех четырех групп, результаты для амплитуды и величины были довольно схожими.В случаях, когда планки погрешностей (± SEM) не видны, ошибка меньше символа.
Есть два дополнительных индикатора, которые подтверждают гипотезу о том, что эта процедура ставит амплитуду ответа под экспериментальный контроль и более точно соответствует естественным условиям. Во-первых, должно быть уменьшение доли испытаний, в которых проводится УЗИ, с почти 1,0 до среднего уровня, что указывает на то, что амплитуда ответа иногда больше, а иногда меньше целевого. A показывает, что действительно доля испытаний, в которых выполнялась УЗИ, уменьшилась по сравнению с тренировкой, что указывает на увеличение количества испытаний, в которых целевая амплитуда была достигнута или превышена. Для этого показателя наблюдалось значительное влияние групп ( F (3,198) = 7,13, p = 0,0047) и тренировочного дня ( F (9,162) = 70,93, p <0,0001 ), но сравнение взаимодействий не было статистически надежным ( F (27,162) = 1.67, p = 0,0545). Во-вторых, должно происходить уменьшение средней ошибки, то есть среднего абсолютного значения разницы между амплитудой отклика и целевой амплитудой. Этот показатель отображается как функция количества тренировочных дней в B . На общее уменьшение средней ошибки указывает значительный эффект тренировочных дней ( F (9 162) = 102,17, p <0,0001). Поскольку начальное значение ошибки различается для четырех групп, неудивительно, что также наблюдался значительный групповой эффект ( F (3,198) = 14.
A показывает, что действительно доля испытаний, в которых выполнялась УЗИ, уменьшилась по сравнению с тренировкой, что указывает на увеличение количества испытаний, в которых целевая амплитуда была достигнута или превышена. Для этого показателя наблюдалось значительное влияние групп ( F (3,198) = 7,13, p = 0,0047) и тренировочного дня ( F (9,162) = 70,93, p <0,0001 ), но сравнение взаимодействий не было статистически надежным ( F (27,162) = 1.67, p = 0,0545). Во-вторых, должно происходить уменьшение средней ошибки, то есть среднего абсолютного значения разницы между амплитудой отклика и целевой амплитудой. Этот показатель отображается как функция количества тренировочных дней в B . На общее уменьшение средней ошибки указывает значительный эффект тренировочных дней ( F (9 162) = 102,17, p <0,0001). Поскольку начальное значение ошибки различается для четырех групп, неудивительно, что также наблюдался значительный групповой эффект ( F (3,198) = 14. 41, p <0,0001) и значительный эффект группы × тренировочные дни ( F (27,162) = 13,6, p <0,0001).
41, p <0,0001) и значительный эффект группы × тренировочные дни ( F (27,162) = 13,6, p <0,0001).
A , B , Уменьшение вероятности доставки в США ( A ) и средней ошибки ( B ) в зависимости от 10 дней обучения для четыре группы амплитуд (2, 3, 4 и 5 мм). Снижение вероятности УЗИ ( A ) указывает на увеличение доли ответов, которые соответствуют целевой амплитуде или превышают ее.Все четыре группы показали значительное снижение вероятности поражения в США, достигнув значений от 21 до 44%. B , Средняя ошибка уменьшилась до аналогичных значений во всех четырех группах.
В целом, эти анализы показывают, что (1) группы, разработанные с разницей в тренировках по амплитуде и величине ответа, (2) доля испытаний, в которых проводилось УЗИ, и средняя ошибка уменьшались за время обучения, и (3) там Нет различий в вероятности ответа для трех групп. Поскольку наиболее простой способ оценить свойства этого обучения – это оценить производительность в стабильном или близком к нему состоянии, далее мы сосредоточили анализ на данных за последние 2 дня обучения.
Поскольку наиболее простой способ оценить свойства этого обучения – это оценить производительность в стабильном или близком к нему состоянии, далее мы сосредоточили анализ на данных за последние 2 дня обучения.
Анализ асимптотической производительности
Для анализа стабильной производительности данные были объединены с 9-го и 10-го дней сбора данных для каждого субъекта. Этот анализ выявил четкие различия в амплитуде и величине CR между четырьмя группами ( A , B ).Отдельные односторонние дисперсионные анализы для амплитуды ( F (3,18) = 19,53, p <0,0001) и величины ( F = 37,25, p <0,0001) подтверждают надежность этих различий. Для измерения величины все шесть парных сравнений (критерий Тьюки, p <0,05) были надежными, а для амплитуды только сравнение 3 и 4 мм не было надежным. В отличие от этих различий, не было достоверных различий в измерении средней ошибки между четырьмя группами ( C ). В качестве финального теста на различия в амплитуде и величине ответов по четырем целевым амплитудным группам были рассчитаны нормализованные совокупные частотные распределения для каждой меры ответа ( G ). Тест Колмогорова – Смирнова ( p <0,005) показал, что все шесть парных сравнений для каждой группы были статистически надежными.
В качестве финального теста на различия в амплитуде и величине ответов по четырем целевым амплитудным группам были рассчитаны нормализованные совокупные частотные распределения для каждой меры ответа ( G ). Тест Колмогорова – Смирнова ( p <0,005) показал, что все шесть парных сравнений для каждой группы были статистически надежными.
Сравнения четырех амплитудных групп при стабильных показателях, взятых для каждого животного с 9-го и 10-го дней. A , Установившаяся амплитуда ответа различалась для четырех групп.Четыре пунктирные линии указывают целевую амплитуду для каждой группы. Все попарные сравнения (критерий Тьюки, p <0,05) были значимыми, за исключением сравнения между группами 3 и 4 мм. B , тот же формат, что и A для данных величины, которые представляют собой среднюю амплитуду всех CR без исключения не-CR. Все попарные сравнения были значимыми (критерий Тьюки, p <0,05). C , Стационарная средняя ошибка (абсолютное значение) существенно не различалась для четырех групп. D , Наблюдалась общая тенденция увеличения доли испытаний, в которых использовались УЗИ, с увеличением амплитуды цели. Единственное парное сравнение, которое было статистически надежным, было 2 против 5 мм. E , Все четыре группы показывают сопоставимые характеристики с точки зрения вероятности CR; между четырьмя группами не было значительных различий. F , Была общая тенденция к увеличению пиковой скорости CR по мере увеличения целевой амплитуды.Единственное достоверное (критерий Тьюки, p <0,05) сравнение было 2 против 5 мм. G , Суммарная доля ответов как функция амплитуды условного ответа (вверху) и величины (внизу) для четырех групп. Каждый CR для каждого животного помещали в бункер (ширина бункера 0,2 мм). Все шесть парных сравнений являются статистически надежными (критерий Колмогорова – Смирнова, p <0,005) для амплитуды, а также для измерения величины (кроме 4 против 5 мм, p <0.
C , Стационарная средняя ошибка (абсолютное значение) существенно не различалась для четырех групп. D , Наблюдалась общая тенденция увеличения доли испытаний, в которых использовались УЗИ, с увеличением амплитуды цели. Единственное парное сравнение, которое было статистически надежным, было 2 против 5 мм. E , Все четыре группы показывают сопоставимые характеристики с точки зрения вероятности CR; между четырьмя группами не было значительных различий. F , Была общая тенденция к увеличению пиковой скорости CR по мере увеличения целевой амплитуды.Единственное достоверное (критерий Тьюки, p <0,05) сравнение было 2 против 5 мм. G , Суммарная доля ответов как функция амплитуды условного ответа (вверху) и величины (внизу) для четырех групп. Каждый CR для каждого животного помещали в бункер (ширина бункера 0,2 мм). Все шесть парных сравнений являются статистически надежными (критерий Колмогорова – Смирнова, p <0,005) для амплитуды, а также для измерения величины (кроме 4 против 5 мм, p <0. 05).
05).
A – C в совокупности предполагают, что, хотя средняя ошибка амплитуды была сопоставима в четырех группах, источники ошибок были разными. Ответы для групп 2 и 3 мм имели тенденцию выходить за цель, тогда как ответы для группы 5 мм имели тенденцию выходить за пределы цели. Эту тенденцию можно вывести из сравнения вероятности в США, показанного в D , которое отражает долю ответов, которые не достигли целевого значения. Несмотря на четкую тенденцию к увеличению этого показателя с целевой амплитудой, различия между группами только приближались к значимости ( F (3,18) = 3.44, p = 0,08). Только группа 2 мм показала значительную тенденцию к превышению целевого значения, потому что отдельные тесты t для каждой группы выявили только достоверное отличие от среднего значения популяции в 50% (равные реакции недостижения и превышения) для группы 2 мм ( t ). (5) = 3,47, p <0,05). Несмотря на разницу в амплитуде и величине ответа, группы показали сопоставимые показатели ( F (3,18) = 1,799, p = 0. 37) с точки зрения вероятности CR ( E ). Наконец, F показывает, что существует также общая тенденция увеличения максимальной скорости КЛ с целевой амплитудой ( F (3,18) = 7,81, p = 0,003). Парные сравнения показали, что различия 2 по сравнению с 4 мм и 2 по сравнению с 5 мм были надежными (критерий Тьюки, p <0,05).
37) с точки зрения вероятности CR ( E ). Наконец, F показывает, что существует также общая тенденция увеличения максимальной скорости КЛ с целевой амплитудой ( F (3,18) = 7,81, p = 0,003). Парные сравнения показали, что различия 2 по сравнению с 4 мм и 2 по сравнению с 5 мм были надежными (критерий Тьюки, p <0,05).
Предыдущие исследования показали, что точное время CR является характерной чертой кондиционирования век (Smith, 1968; Smith et al., 1969; Милленсон и др., 1977; Маук и Руис, 1992). Время ожидания и время нарастания ответов систематически меняются в зависимости от межстимульного интервала, так что ответы имеют тенденцию достигать пика около времени доставки УЗИ. Мы исследовали различные меры времени ответа для четырех целевых амплитудных групп и не обнаружили достоверных различий во времени ответа ( A ). Ни время ожидания начала, ни время ожидания критерия, время ожидания максимальной скорости, ни время ожидания ответа пика не показали статистически достоверных различий между группами. Когда ответы были разделены в зависимости от того, превышают ли они цель (испытания в США) или ниже цели (испытания в США), наблюдались значительные различия во времени ответов во всех четырех группах ( B ). Отдельные двухсторонние смешанные ANOVA для задержки начала и задержки до максимальной скорости показали только значимое влияние типа ответа (не в США по сравнению с США; начало, F (1,18) = 100,6, p < 0,0001; пиковая скорость, F = 49.04, p <0,0001). Этот анализ показывает, что в целом ответы, которые соответствовали целевой амплитуде или превышали ее, имели более короткие задержки, чем ответы, которые не соответствовали целевой амплитуде.
Когда ответы были разделены в зависимости от того, превышают ли они цель (испытания в США) или ниже цели (испытания в США), наблюдались значительные различия во времени ответов во всех четырех группах ( B ). Отдельные двухсторонние смешанные ANOVA для задержки начала и задержки до максимальной скорости показали только значимое влияние типа ответа (не в США по сравнению с США; начало, F (1,18) = 100,6, p < 0,0001; пиковая скорость, F = 49.04, p <0,0001). Этот анализ показывает, что в целом ответы, которые соответствовали целевой амплитуде или превышали ее, имели более короткие задержки, чем ответы, которые не соответствовали целевой амплитуде.
Меры времени отклика по сравнению с четырьмя группами амплитуды. A , Четыре графика показывают задержку до начала, задержку до критерия, задержку до пиковой скорости и задержку до пиковой амплитуды (для испытаний за пределами США). Статистически достоверной разницы между группами по этим показателям ответа не было. B , Разница во времени испытаний в США и за пределами США. Раздельный анализ исследований в УЗИ (амплитуда ниже целевого) и неамериканских исследований (амплитуда на уровне или выше) выявил общую тенденцию к более быстрым и коротким задержкам ответов в неамериканских исследованиях. Все восемь сравнений (4 группы × 2 меры ответа) были статистически надежными (парный тест t , p <0,01).
B , Разница во времени испытаний в США и за пределами США. Раздельный анализ исследований в УЗИ (амплитуда ниже целевого) и неамериканских исследований (амплитуда на уровне или выше) выявил общую тенденцию к более быстрым и коротким задержкам ответов в неамериканских исследованиях. Все восемь сравнений (4 группы × 2 меры ответа) были статистически надежными (парный тест t , p <0,01).
Примеры необработанных ответов от одного испытуемого из каждой амплитудной группы (день 10 в каждом случае) показаны в.Для каждого субъекта ответы, которые соответствовали целевой амплитуде или превышали ее, показаны в левом столбце, а те, которые не соответствовали целевой амплитуде (и, следовательно, были доставлены УЗИ), показаны в правом столбце. Эти данные показывают общую тенденцию реакции к перерегулированию для меньшей амплитуды цели и к недолету для цели 5 мм. Вместе эти анализы демонстрируют, что процедура тренировки целевой амплитуды может использоваться для получения разумного экспериментального контроля над амплитудой ответа при кондиционировании век, что средняя ошибка амплитуды сопоставима для целей от 2 до 5 мм, и что эти различия возникают на сопоставимых уровнях CR. и сопоставимые статистические данные о сроках проведения CR.
и сопоставимые статистические данные о сроках проведения CR.
Пример необработанных ответов от четырех животных, по одному от каждой группы амплитуд, взятых с 10-го дня обучения. Каждая развертка представляет собой отдельную реакцию века (первая проба впереди), причем черная область каждой развертки указывает продолжительность CS. Ответы были разделены на испытания, проводимые за пределами США (левый столбец) и испытания в США (правый столбец). Последовательность испытаний каждого животного в США и за пределами США очевидна по интервалу между испытаниями. Обратите внимание на общую тенденцию к большему количеству испытаний без УЗИ с малыми амплитудами цели и к большему количеству испытаний УЗИ с большими амплитудами цели.Компонент рефлекторного ответа каждого испытания виден в светло-серой области испытаний «With-US».
Амплитуда отклика после повторного обнаружения новой цели
Практическая и концептуальная важность тренировки амплитуды целевой будет уменьшена, если амплитуды отклика цели не могут быть получены и повторно получены в соответствии с планом эксперимента. Поэтому мы тестировали каждое животное, используя новую целевую амплитуду, отличную от начальной. После 10 дней обучения до начальной целевой амплитуды каждый субъект был обучен в течение 10 дополнительных дней с использованием новой целевой амплитуды.Новые целевые амплитуды для групп 2, 3, 4 и 5 мм, соответственно, составляли 4, 5, 2 и 3 мм. показывает сравнение амплитуд ответа для этих четырех групп для начальной и новой целевой амплитуд. Как и выше, эти данные включали объединенное среднее значение 9 и 10 дней для каждого субъекта. Мы начали с того, что спросили, есть ли различия между получением и повторным обнаружением для каждой из четырех целевых амплитуд (левый график). Это сравнение между субъектами сравнивает каждую цель, когда она использовалась при захвате, по сравнению с повторным захватом (т.е., получение от группы 2 мм и повторное обнаружение на цели 2 мм, первоначально группы 4 мм). Двусторонний дисперсионный анализ выявил только основной эффект целевой амплитуды ( F (3,36) = 37,06, p <0,0001), эффекты захвата по сравнению с повторным захватом и взаимодействия не были значительными, и не было надежных различия в амплитудах, полученных во время захвата, по сравнению с повторным обнаружением новой цели.
Поэтому мы тестировали каждое животное, используя новую целевую амплитуду, отличную от начальной. После 10 дней обучения до начальной целевой амплитуды каждый субъект был обучен в течение 10 дополнительных дней с использованием новой целевой амплитуды.Новые целевые амплитуды для групп 2, 3, 4 и 5 мм, соответственно, составляли 4, 5, 2 и 3 мм. показывает сравнение амплитуд ответа для этих четырех групп для начальной и новой целевой амплитуд. Как и выше, эти данные включали объединенное среднее значение 9 и 10 дней для каждого субъекта. Мы начали с того, что спросили, есть ли различия между получением и повторным обнаружением для каждой из четырех целевых амплитуд (левый график). Это сравнение между субъектами сравнивает каждую цель, когда она использовалась при захвате, по сравнению с повторным захватом (т.е., получение от группы 2 мм и повторное обнаружение на цели 2 мм, первоначально группы 4 мм). Двусторонний дисперсионный анализ выявил только основной эффект целевой амплитуды ( F (3,36) = 37,06, p <0,0001), эффекты захвата по сравнению с повторным захватом и взаимодействия не были значительными, и не было надежных различия в амплитудах, полученных во время захвата, по сравнению с повторным обнаружением новой цели. Правая часть показывает те же данные, построенные заново, чтобы выделить сравнения амплитуд, полученных для сбора и повторного сбора данных в одних и тех же группах.Четыре отдельных парных теста t ( p <0,01) показали, что для каждой группы амплитуда ответов, полученных при обнаружении одной цели, отличалась от тех, которые были получены при повторном обнаружении на новую цель. Эти анализы показывают, что как получение первой целевой амплитуды, так и повторное обнаружение новой целевой амплитуды вызвали значительную разницу в амплитуде ответа для каждой целевой амплитудной группы и что не было никаких различий в этих тенденциях между захватом и повторным захватом.
Правая часть показывает те же данные, построенные заново, чтобы выделить сравнения амплитуд, полученных для сбора и повторного сбора данных в одних и тех же группах.Четыре отдельных парных теста t ( p <0,01) показали, что для каждой группы амплитуда ответов, полученных при обнаружении одной цели, отличалась от тех, которые были получены при повторном обнаружении на новую цель. Эти анализы показывают, что как получение первой целевой амплитуды, так и повторное обнаружение новой целевой амплитуды вызвали значительную разницу в амплитуде ответа для каждой целевой амплитудной группы и что не было никаких различий в этих тенденциях между захватом и повторным захватом.
Сравнение устойчивых амплитуд ответа после первоначального сбора данных (темно-серый; те же данные, что и A ) и у тех же животных после 10 дней повторного обнаружения с новой целевой амплитудой. Начальная амплитуда цели и амплитуда цели повторного обнаружения были, соответственно, для четырех групп (от 2 до 4 мм, от 3 до 5 мм, от 4 до 2 мм и от 5 до 3 мм). Как и в случае, данные получены из среднего значения для каждого животного за 9-й и 10-й дни захвата (и повторного захвата). Слева: сравнение идентичных целевых амплитуд для захвата и повторного захвата.Статистически достоверных различий для каждой целевой амплитуды между данными сбора и повторного обнаружения не было ( t тестов). Целевая амплитуда обозначена пунктирными линиями. Справа: сравнение данных приобретения и повторного приобретения для одних и тех же групп животных. Различные целевые амплитуды захвата и повторного захвата для каждой группы снова обозначены пунктирными линиями. Парные тесты t ( p <0,01) показали статистически достоверные различия между амплитудами сбора и повторного обнаружения для каждой группы.
Как и в случае, данные получены из среднего значения для каждого животного за 9-й и 10-й дни захвата (и повторного захвата). Слева: сравнение идентичных целевых амплитуд для захвата и повторного захвата.Статистически достоверных различий для каждой целевой амплитуды между данными сбора и повторного обнаружения не было ( t тестов). Целевая амплитуда обозначена пунктирными линиями. Справа: сравнение данных приобретения и повторного приобретения для одних и тех же групп животных. Различные целевые амплитуды захвата и повторного захвата для каждой группы снова обозначены пунктирными линиями. Парные тесты t ( p <0,01) показали статистически достоверные различия между амплитудами сбора и повторного обнаружения для каждой группы.
Стимуляция мшистых волокон в качестве CS
Значительные доказательства указывают на то, что приобретение и экспрессия CR во время формирования павлового (отложенного) века опосредуется пластичностью мозжечка. Поскольку обусловленность целевой амплитуды, строго говоря, не является павловской, мы провели два дополнительных эксперимента, чтобы проверить возможность того, что обучение в этих условиях опосредуется нецеребеллярными механизмами. Способность заменять стимуляцию мшистых волокон тонусом CS – одно из наиболее убедительных доказательств, подтверждающих, что мозжечковые механизмы являются основой кондиционирования век.Таким образом, мы сравнили эффекты тренировки с целевой амплитудой 3 мм, используя тон CS в одной группе и стимуляцию мшистых волокон в качестве CS во второй группе. В остальном процедуры были идентичны процедурам, использованным выше для группы 3 мм. показывает результаты за 10 дней тренировок для этих двух групп. A – D сравнивает эти две группы за 10 дней обучения, а E показывает сравнения в установившемся состоянии, как в предыдущем эксперименте. Отдельные двухфакторные смешанные ANOVA для четырех мер реакции показали только основные эффекты тренировочных дней (амплитуда, F (9,54) = 30.54; ошибка, F = 17,71; Вероятность CR, F = 29,09; Вероятность США – 16,62; p <0,0001 во всех случаях). Ни один из основных эффектов группы или эффектов взаимодействия не был значимым, равно как и сравнение тона и мшистого волокна ( E ) для восьми показателей реакции, которые были проанализированы для данных устойчивого состояния (дни 9 и 10, t тесты, p > 0,05). Эти данные исключают факторы, расположенные выше мозжечка, как способствующие приобретению амплитуды ответа, и подтверждают гипотезу о том, что кондиционирование век к целевой амплитуде опосредуется мозжечком.
Способность заменять стимуляцию мшистых волокон тонусом CS – одно из наиболее убедительных доказательств, подтверждающих, что мозжечковые механизмы являются основой кондиционирования век.Таким образом, мы сравнили эффекты тренировки с целевой амплитудой 3 мм, используя тон CS в одной группе и стимуляцию мшистых волокон в качестве CS во второй группе. В остальном процедуры были идентичны процедурам, использованным выше для группы 3 мм. показывает результаты за 10 дней тренировок для этих двух групп. A – D сравнивает эти две группы за 10 дней обучения, а E показывает сравнения в установившемся состоянии, как в предыдущем эксперименте. Отдельные двухфакторные смешанные ANOVA для четырех мер реакции показали только основные эффекты тренировочных дней (амплитуда, F (9,54) = 30.54; ошибка, F = 17,71; Вероятность CR, F = 29,09; Вероятность США – 16,62; p <0,0001 во всех случаях). Ни один из основных эффектов группы или эффектов взаимодействия не был значимым, равно как и сравнение тона и мшистого волокна ( E ) для восьми показателей реакции, которые были проанализированы для данных устойчивого состояния (дни 9 и 10, t тесты, p > 0,05). Эти данные исключают факторы, расположенные выше мозжечка, как способствующие приобретению амплитуды ответа, и подтверждают гипотезу о том, что кондиционирование век к целевой амплитуде опосредуется мозжечком.
Получение целевой амплитуды (3 мм) для животных, обученных с помощью тонального CS и стимуляции мшистыми волокнами в качестве CS. A , Амплитуда CR за 10 дней обучения. B , Средняя ошибка (абсолютное значение) за 10 дней обучения. MF, Мшистое волокно. C , D , вероятность CR ( C ) и вероятность США ( D ) за 10 дней обучения. E , Сравнение между группами CS тональной стимуляции и CS со стимуляцией мшистыми волокнами для различных мер реакции, взятых с 9-го и 10-го дней тренировки.Ни одно из шести сравнений групп не было статистически надежным (тест t , p > 0,05).
Обратимая инактивация мозжечка
В качестве дополнительной проверки гипотезы о том, что мозжечок необходим для кондиционирования до заданной амплитуды, экспрессия CR была протестирована на четырех животных, обученных тону, и четырех животных, обученных мшистым волокнам, до и после инактивации. мозжечкового выхода через инфузию мусцимола, агониста ГАМК, в переднее межположительное ядро ().Субъекты в обеих группах тестировались в отдельные дни с инфузиями носителя или инфузиями мусцимола (порядок уравновешивался для каждой группы). График средних показателей по группе (в центре) показывает, что для обеих групп не было достоверных изменений после инфузии носителя, тогда как инфузии мусцимола отменяли экспрессию CR. Необработанные данные для одного субъекта стимуляции мшистых волокон для инфузий носителя (слева) и мусцимола (справа) показаны вверху. Пример канюли (слева) и стимулирующих электродов от того же пациента показаны внизу.
Необходимость мозжечка для выражения приобретенных ответов на заданную амплитуду 3 мм. Две группы, одна из которых использовала тональный CS, а другая, в которой стимуляция волокон использовалась в качестве CS, были протестированы в отдельные дни либо с инфузией носителя, либо с инфузией мусцимола, агониста ГАМК. На двух графиках показаны средние показатели по группе отсутствия эффекта от инфузии носителя (слева) и отмены CR с помощью инфузии мусцимола (справа). Время настаивания в каждом случае указано черной стрелкой.Примеры исходных данных от одного животного показаны выше: следы слева – это инфузия носителя, а следы справа – инфузия мусцимола. В каждом случае ответы до инфузии находятся впереди (первое испытание впереди), а реакции после инфузии – сзади. На двух нижних фотографиях показано размещение инфузионной канюли (слева) и стимулирующих электродов из мшистых волокон (справа) для животного, исходные данные которого показаны выше. Этому животному были имплантированы два электрода из покрытых мхом волокон; только один был использован для этого эксперимента.MF, Мшистые волокна.
Две группы, одна из которых использовала тональный CS, а другая, в которой стимуляция волокон использовалась в качестве CS, были протестированы в отдельные дни либо с инфузией носителя, либо с инфузией мусцимола, агониста ГАМК. На двух графиках показаны средние показатели по группе отсутствия эффекта от инфузии носителя (слева) и отмены CR с помощью инфузии мусцимола (справа). Время настаивания в каждом случае указано черной стрелкой.Примеры исходных данных от одного животного показаны выше: следы слева – это инфузия носителя, а следы справа – инфузия мусцимола. В каждом случае ответы до инфузии находятся впереди (первое испытание впереди), а реакции после инфузии – сзади. На двух нижних фотографиях показано размещение инфузионной канюли (слева) и стимулирующих электродов из мшистых волокон (справа) для животного, исходные данные которого показаны выше. Этому животному были имплантированы два электрода из покрытых мхом волокон; только один был использован для этого эксперимента.MF, Мшистые волокна.
В совокупности исследования стимуляции мшистых волокон и инактивации глубоких ядер показывают (1), что мозжечок необходим для экспрессии CR, продуцируемых кондиционированием века до целевой амплитуды (результаты инфузии), и (2) что механизмы, ответственные за приобретение и экспрессия целевой амплитуды ответа не находятся в пути CR выше мозжечка (результаты стимуляции мшистыми волокнами).
Обсуждение
Мы адаптировали традиционные процедуры кондиционирования павловских век, чтобы ввод ошибки, определяемый США, зависел от амплитуды CR.Пропустив УЗИ в исследованиях, в которых CR соответствует или превышает заданную целевую амплитуду (и представив УЗИ в противном случае), мы показали, что результирующие условные реакции век имеют тенденцию к целевой амплитуде. Это верно как для получения исходной целевой амплитуды, так и для повторного обнаружения с использованием другой цели. Несмотря на то, что наблюдалась тенденция к перерегулированию при малых амплитудах цели и к недолету с наибольшей целью, средняя абсолютная ошибка не различалась для групп, обученных для целевых амплитуд от 2 до 5 мм. Эти данные показывают, что, адаптируя процедуры кондиционирования век к естественным условиям обучения мозжечка, амплитуду CR можно поставить под экспериментальный контроль. Способность устранять эти CR с помощью инактивации мозжечка и способность заменять стимуляцию мшистых волокон тональным CS подтверждают гипотезу о том, что, как и в случае кондиционирования века в форме павлова, кондиционирование века до целевой амплитуды опосредуется мозжечком. Эти результаты формируют основу для систематических и контролируемых исследований того, как мозжечок обучается, сохраняет и выражает надлежащую амплитуду откликов, которые он адаптирует.
Эти данные показывают, что, адаптируя процедуры кондиционирования век к естественным условиям обучения мозжечка, амплитуду CR можно поставить под экспериментальный контроль. Способность устранять эти CR с помощью инактивации мозжечка и способность заменять стимуляцию мшистых волокон тональным CS подтверждают гипотезу о том, что, как и в случае кондиционирования века в форме павлова, кондиционирование века до целевой амплитуды опосредуется мозжечком. Эти результаты формируют основу для систематических и контролируемых исследований того, как мозжечок обучается, сохраняет и выражает надлежащую амплитуду откликов, которые он адаптирует.
Значение для кондиционирования век и мозжечка
Необходимость мозжечка для кондиционирования века с задержкой павлова подтверждается большим количеством данных. Настоящие результаты показывают, что непредвиденные обстоятельства Павлова, при которых доставка УЗИ не зависит от CR, не являются особенностью, которая делает мозжечок необходимым. Поскольку, строго говоря, процедуры тренировки до целевой амплитуды включают в себя оперантные непредвиденные обстоятельства, необходимость мозжечка в кондиционировании век связана с природой стимулов и системой реакции, а не с природой непредвиденных обстоятельств, которые управляют представлением усиливающий стимул.
Настоящие данные также обращаются к распространенному заблуждению о кондиционировании век, а именно, что это рефлекторное кондиционирование или кондиционирование моргания глаз. Павлов выступил с идеей, что классическое кондиционирование включает в себя пути, активируемые CS, обретающие способность активировать путь, который опосредует рефлекторный ответ (Павлов, 1927). Нервные пути, которые, как известно, опосредуют экспрессию CR и рефлекторные мигания, вызываемые США, не согласуются с гипотезой Павлова. Рефлекторные мигания, вызываемые первичными афферентами, проходящими через верхний глазничный нерв, опосредуются двумя путями (Pellegrini et al., 1995). Первичные афференты проецируются как на каудально-межполярную пограничную область спинномозгового ядра тройничного нерва, так и на дорсальный рог, сегмент С1 спинного мозга. Обе эти области проецируются в область orbicularis oculi лицевого ядра. Эксперименты с поражением показали, что спинальный тройничный путь опосредует начальный компонент R1 рефлекторных морганий, а путь спинного мозга опосредует более длинный латентный компонент R2 (Pelligrini et al. , 1995). Путь CR, выступающий из переднего межположительного ядра через красное ядро, по-видимому, соединяется с этим путем в двигательных или премоторных нейронах лицевого ядра.Следовательно, рефлекторные мигания и CRs опосредуются совершенно разными нервными путями и, как можно ожидать, будут проявлять совершенно разные свойства, как было четко установлено в исследованиях на людях (Schade Powers et al., 2010). Поэтому условные реакции век более правильно охарактеризовать как «реакции век», чем «моргания глаз», и настоящие данные демонстрируют, что амплитуда этих ответов век, как и большинство других реакций скелетных мышц, может быть изучена и опосредована обучением мозжечка.
, 1995). Путь CR, выступающий из переднего межположительного ядра через красное ядро, по-видимому, соединяется с этим путем в двигательных или премоторных нейронах лицевого ядра.Следовательно, рефлекторные мигания и CRs опосредуются совершенно разными нервными путями и, как можно ожидать, будут проявлять совершенно разные свойства, как было четко установлено в исследованиях на людях (Schade Powers et al., 2010). Поэтому условные реакции век более правильно охарактеризовать как «реакции век», чем «моргания глаз», и настоящие данные демонстрируют, что амплитуда этих ответов век, как и большинство других реакций скелетных мышц, может быть изучена и опосредована обучением мозжечка.
Значение для обучения мозжечка усилению или амплитуде
Вопрос о том, как и где мозжечок обучается и сохраняет усиление или амплитуду ответа, взаимосвязан с вопросом о месте (ах) пластичности, ответственных за обучение мозжечка. На этот вопрос лучше всего ответить с помощью исследований, включающих анализ адаптации VOR (Raymond et al. , 1996; Boyden et al., 2004). Первоначальные усилия в этой области вызвали серьезные дискуссии о том, происходит ли пластичность в коре мозжечка или в вестибулярном ядре, чьи связи и схема параллельны глубоким ядрам мозжечка (Lisberger and Sejnowski, 1992, 1993; Ito, 1993).Более поздние теории предполагают участие пластичности как в коре мозжечка, так и в глубоком ядре (Perrett et al., 1993; Raymond et al., 1996; Mauk and Donegan, 1997). Когда задействовано более одного сайта пластичности, становится важным вопрос об относительном вкладе этих двух сайтов. Относительный вклад коры мозжечка и механизмов пластичности глубокого ядра мозжечка в усиление или амплитуду ответа остается нерешенным, и есть указания на то, что относительный вклад может варьироваться сложным образом в зависимости от обстоятельств (Mauk, 1997).Кондиционирование век до целевой амплитуды обеспечивает способ решения этих вопросов с помощью относительно простой и экспериментально управляемой системы, которая поддается параметрическому поведенческому анализу, обратимым повреждениям, in vivo записи и даже крупномасштабному компьютерному моделированию.
, 1996; Boyden et al., 2004). Первоначальные усилия в этой области вызвали серьезные дискуссии о том, происходит ли пластичность в коре мозжечка или в вестибулярном ядре, чьи связи и схема параллельны глубоким ядрам мозжечка (Lisberger and Sejnowski, 1992, 1993; Ito, 1993).Более поздние теории предполагают участие пластичности как в коре мозжечка, так и в глубоком ядре (Perrett et al., 1993; Raymond et al., 1996; Mauk and Donegan, 1997). Когда задействовано более одного сайта пластичности, становится важным вопрос об относительном вкладе этих двух сайтов. Относительный вклад коры мозжечка и механизмов пластичности глубокого ядра мозжечка в усиление или амплитуду ответа остается нерешенным, и есть указания на то, что относительный вклад может варьироваться сложным образом в зависимости от обстоятельств (Mauk, 1997).Кондиционирование век до целевой амплитуды обеспечивает способ решения этих вопросов с помощью относительно простой и экспериментально управляемой системы, которая поддается параметрическому поведенческому анализу, обратимым повреждениям, in vivo записи и даже крупномасштабному компьютерному моделированию.
Добавляя, сколько и когда
Abstract
Концептуальные и практические преимущества кондиционирования павловых век облегчают анализ вычислений и обучения мозжечка.Тем не менее, процедуры кондиционирования век во многом нереальны. Сигнал ошибки для оливоцеребеллярной системы не уменьшается, поскольку обучение адаптирует амплитуду или усиление ответа. Это по своей сути ограничивает полезность кондиционирования век для исследований, изучающих, как механизмы обучения мозжечка приобретают и сохраняют амплитуду адаптивного ответа. Мы сообщаем о разработке и описании процедуры обучения, в которой амплитуда условной реакции ставится под экспериментальный контроль с непредвиденными обстоятельствами, которые более близки к естественным условиям.В этой процедуре доставка безусловного стимула (US) зависит от амплитуды условного ответа: US доставляется для ответов, которые не достигают заданной целевой амплитуды, и опускается для ответов, которые соответствуют или превышают цель. Мы обнаружили, что кролики, обученные либо тональной стимуляции, либо стимуляции мшистыми волокнами в качестве условного стимула, усваивают ответы, которые достигают целевых амплитуд в диапазоне от 2 до 5 мм. Инактивация межположительного ядра с помощью инфузий мусцимола устраняет эти условные реакции, указывая на то, что участие мозжечка в кондиционировании век не связано явно с использованием процедур Павлова.Вместе с предыдущими исследованиями эти данные предполагают, что амплитуда ответа изучается и кодируется в мозжечке во время кондиционирования век. Таким образом, эти результаты обеспечивают основу для систематических и контролируемых исследований мозжечковых механизмов, которые обучаются и кодируют правильную амплитуду адаптивных движений.
Мы обнаружили, что кролики, обученные либо тональной стимуляции, либо стимуляции мшистыми волокнами в качестве условного стимула, усваивают ответы, которые достигают целевых амплитуд в диапазоне от 2 до 5 мм. Инактивация межположительного ядра с помощью инфузий мусцимола устраняет эти условные реакции, указывая на то, что участие мозжечка в кондиционировании век не связано явно с использованием процедур Павлова.Вместе с предыдущими исследованиями эти данные предполагают, что амплитуда ответа изучается и кодируется в мозжечке во время кондиционирования век. Таким образом, эти результаты обеспечивают основу для систематических и контролируемых исследований мозжечковых механизмов, которые обучаются и кодируют правильную амплитуду адаптивных движений.
Введение
Павловское кондиционирование века облегчило анализ обучения и вычислений мозжечка (McCormick and Thompson, 1984; Perrett et al., 1993; Маук и Донеган, 1997; Медина и Маук, 1999; Hansel et al., 2001; Medina et al., 2001, 2002), в основном из-за тесного соответствия между тренировочными стимулами и входными сигналами мозжечка, а также между выходной мощностью мозжечка и поведенческими реакциями (McCormick and Thompson, 1984; Mauk et al. , 1986; Steinmetz et al. , 1986; Mauk, Donegan, 1997; Hesslow et al., 1999). Идея Павлова (1927) о том, что условное кондиционирование – это «рефлексивное обучение», при котором условный стимул (КС) приобретает способность активировать рефлекторный путь, запускаемый подкрепляющим стимулом, является неправильным представлением, обычно применяемым к кондиционированию век.В самом деле, неправильное название «кондиционирование моргания», по-видимому, происходит из этого заблуждения. Данные, противоречащие этой идее, включают известные схемы кондиционирования век, которые ясно показывают, что путь CS не связан с рефлекторным путем через обучение (Mauk and Donegan, 1997; Thompson et al., 1998), из доказательств того, что условные реакции век не являются морганиями (Schade Powers et al., 2010), а из простого поведенческого наблюдения, что во время приобретения амплитуда условной реакции не является однозначной.С тренировкой амплитуда условной реакции монотонно растет от едва заметного до максимального закрытия век.
, 1986; Steinmetz et al. , 1986; Mauk, Donegan, 1997; Hesslow et al., 1999). Идея Павлова (1927) о том, что условное кондиционирование – это «рефлексивное обучение», при котором условный стимул (КС) приобретает способность активировать рефлекторный путь, запускаемый подкрепляющим стимулом, является неправильным представлением, обычно применяемым к кондиционированию век.В самом деле, неправильное название «кондиционирование моргания», по-видимому, происходит из этого заблуждения. Данные, противоречащие этой идее, включают известные схемы кондиционирования век, которые ясно показывают, что путь CS не связан с рефлекторным путем через обучение (Mauk and Donegan, 1997; Thompson et al., 1998), из доказательств того, что условные реакции век не являются морганиями (Schade Powers et al., 2010), а из простого поведенческого наблюдения, что во время приобретения амплитуда условной реакции не является однозначной.С тренировкой амплитуда условной реакции монотонно растет от едва заметного до максимального закрытия век.
Тем не менее, стандартные тренировочные процедуры кондиционирования век не позволяют экспериментально контролировать амплитуду ответа. Для более естественных реакций, таких как адаптация вестибулоокулярного рефлекса (VOR), вход ошибки в оливо-церебеллярную систему (т. Е. Входной сигнал восходящего волокна в мозжечок) уменьшается по мере того, как обучение адаптирует усиление или амплитуду ответа (Gilbert and Thach, 1977). ; Раймонд и Лисбергер, 1997, 1998).Поскольку вероятность сигнала ошибки в оливоцеребеллярной системе [через предъявление безусловного стимула (УЗ)] не уменьшается по мере роста амплитуды условного ответа, процедуры Павлова по своей природе способствуют условным ответам (КО) с максимальной амплитудой (5–6 мм для кроликов). ). Даже при воздушной затяжке, такой как УЗИ, CR (расширение мигательной перепонки) не может полностью устранить активацию УЗ-лазейных волокон, потому что внешние веки обычно ограничены, а сама мигательная перепонка весьма чувствительна.По этим причинам кондиционирование век не было полезным для анализа мозжечковых механизмов, участвующих в обучении и кодировании амплитуды ответа (то есть усиления ответа).
Мы сообщаем о разработке и описании процедуры тренировки век, которая более точно имитирует естественные требования к обучению мозжечка и ставит амплитуду ответа под экспериментальный контроль. В этой процедуре УЗИ зависит от амплитуды условного ответа, так же как ошибочные воздействия на мозжечок зависят от усиления ответа.Когда амплитуда ответа соответствует заранее заданной целевой амплитуде или превышает ее, УЗИ не выполняется (см.). Мы демонстрируем, что амплитуда ответа приближается к целевым амплитудам в диапазоне от 2 до 5 мм, со значительными групповыми различиями при анализе после 10 тренировок. Участие мозжечка в этой обучающей задаче подтверждается двумя наблюдениями: (1) амплитуда ответа может быть определена, когда стимуляция мшистых волокон заменяет тон, и (2) инактивация мозжечкового выхода путем вливания мусцимола в межположительное ядро устраняет экспрессию CR, что позволяет предположить что мозжечковое обучение отвечает не только за то, нужно ли и когда реагировать, но и за то, насколько сильно реагировать. Эти данные показывают, что эта новая тренировочная процедура представляет собой управляемую экспериментальную парадигму для изучения мозжечковых механизмов, которые обучаются и кодируют амплитуду ответа.
Эти данные показывают, что эта новая тренировочная процедура представляет собой управляемую экспериментальную парадигму для изучения мозжечковых механизмов, которые обучаются и кодируют амплитуду ответа.
Схематическое изображение тренировочных процедур, используемых для тренировки реакции век на заданную амплитуду. Каждое тренировочное испытание включало предъявление тонального сигнала CS и запланированную доставку ультразвуковой стимуляции глаза с межстимульным интервалом 500 мс. Фактическая доставка УЗИ зависела от амплитуды ответа путем мониторинга амплитуды ответа в реальном времени во время CS.За десять миллисекунд до запланированной доставки США (время принятия решения в США) обучающее программное обеспечение было запрограммировано таким образом, чтобы определять, будут ли доставлены УЗИ: для ответов, которые соответствовали или превышали заданную амплитуду, доставлялись УЗИ; для ответов, которые не достигли целевой амплитуды, доставлялся УЗИ.
Результаты
Получение целевой амплитуды отклика
В естественных условиях, таких как адаптация VOR, преследования или саккад, вызванное ошибкой возбуждающее воздействие на вход восходящего волокна в мозжечок уменьшается по мере того, как усиление отклика приближается к правильному значению (Гилберт и Тач, 1977; Раймонд и Лисбергер, 1997, 1998). Наша процедура обучения предназначена для более точного приближения к этим естественным условиям обучения путем связывания УЗ-презентации с амплитудой ответа относительно целевой амплитуды. Для этого мы тренировали четыре группы животных в течение 10 дней, каждая группа подвергалась одной из четырех целевых амплитуд (2, 3, 4 или 5 мм). Двадцать четыре кролика были случайным образом отнесены к одной из этих групп, при этом два животных из группы 5 мм в конечном итоге были исключены из-за технических трудностей. Для каждого ежедневного сеанса УЗИ не проводилось в испытаниях, в которых ответ век достигал целевой амплитуды в пределах 10 мс после введения УЗИ.УЗИ проводилось в испытаниях, в которых реакция века не достигала целевой амплитуды.
Наша процедура обучения предназначена для более точного приближения к этим естественным условиям обучения путем связывания УЗ-презентации с амплитудой ответа относительно целевой амплитуды. Для этого мы тренировали четыре группы животных в течение 10 дней, каждая группа подвергалась одной из четырех целевых амплитуд (2, 3, 4 или 5 мм). Двадцать четыре кролика были случайным образом отнесены к одной из этих групп, при этом два животных из группы 5 мм в конечном итоге были исключены из-за технических трудностей. Для каждого ежедневного сеанса УЗИ не проводилось в испытаниях, в которых ответ век достигал целевой амплитуды в пределах 10 мс после введения УЗИ.УЗИ проводилось в испытаниях, в которых реакция века не достигала целевой амплитуды.
Как показано на, животные в четырех группах приобрели устойчивые CR, и для каждой группы средняя амплитуда / величина ответов приближается к заданной цели. Для измерения амплитуды были включены все ответы, тогда как при измерении амплитуды были исключены все не-CR (те, у которых амплитуда <0,3 мм).![]() Каждый из отдельных двухсторонних смешанных дисперсионных анализов (один для амплитуды и один для величины) показал значительную группу (амплитуда, F (3,198) = 7.43, p = 0,0039; величина, F = 11,18, p = 0,00013), тренировочный день (амплитуда, F (9,162) = 126,4, p <0,0001; величина, F = 101,21, p <0,0001 ) и взаимодействие группа × день (амплитуда, F (27,162) = 2,99, p <0,0001; величина, F = 3,66, p <0,0001). Эти анализы показывают, что животные учились, что между группами были различия в амплитуде и величине ответов, и что эти различия развивались в процессе обучения.Несмотря на эти различия в амплитуде и величине ответа, группы были сопоставимы по доле испытаний, в которых произошел полный ответ ( C ). Для этих данных двусторонний смешанный дисперсионный анализ показал значительный эффект дня тренировки ( F (9,162) = 123,8, p <0,0001), но не было никаких эффектов взаимодействия группы или группы × день тренировки ( p > 0,05).
Каждый из отдельных двухсторонних смешанных дисперсионных анализов (один для амплитуды и один для величины) показал значительную группу (амплитуда, F (3,198) = 7.43, p = 0,0039; величина, F = 11,18, p = 0,00013), тренировочный день (амплитуда, F (9,162) = 126,4, p <0,0001; величина, F = 101,21, p <0,0001 ) и взаимодействие группа × день (амплитуда, F (27,162) = 2,99, p <0,0001; величина, F = 3,66, p <0,0001). Эти анализы показывают, что животные учились, что между группами были различия в амплитуде и величине ответов, и что эти различия развивались в процессе обучения.Несмотря на эти различия в амплитуде и величине ответа, группы были сопоставимы по доле испытаний, в которых произошел полный ответ ( C ). Для этих данных двусторонний смешанный дисперсионный анализ показал значительный эффект дня тренировки ( F (9,162) = 123,8, p <0,0001), но не было никаких эффектов взаимодействия группы или группы × день тренировки ( p > 0,05).
Получение условных ответов век за 10 дней обучения для четырех групп, которым были назначены разные целевые амплитуды (2, 3, 4 и 5 мм). A , Амплитуда условных ответов век за 10 дней тренировки показывает получение разных амплитуд ответов для четырех групп. B , Аналогичные результаты были получены для измерения величины отклика, которая исключила ответы, которые не достигли критерия 0,3 мм, чтобы считаться CR. C , Поскольку вероятность ответа была устойчивой для всех четырех групп, результаты для амплитуды и величины были довольно схожими.В случаях, когда планки погрешностей (± SEM) не видны, ошибка меньше символа.
Есть два дополнительных индикатора, которые подтверждают гипотезу о том, что эта процедура ставит амплитуду ответа под экспериментальный контроль и более точно соответствует естественным условиям. Во-первых, должно быть уменьшение доли испытаний, в которых проводится УЗИ, с почти 1,0 до среднего уровня, что указывает на то, что амплитуда ответа иногда больше, а иногда меньше целевого. A показывает, что действительно доля испытаний, в которых выполнялась УЗИ, уменьшилась по сравнению с тренировкой, что указывает на увеличение количества испытаний, в которых целевая амплитуда была достигнута или превышена. Для этого показателя наблюдалось значительное влияние групп ( F (3,198) = 7,13, p = 0,0047) и тренировочного дня ( F (9,162) = 70,93, p <0,0001 ), но сравнение взаимодействий не было статистически надежным ( F (27,162) = 1.67, p = 0,0545). Во-вторых, должно происходить уменьшение средней ошибки, то есть среднего абсолютного значения разницы между амплитудой отклика и целевой амплитудой. Этот показатель отображается как функция количества тренировочных дней в B . На общее уменьшение средней ошибки указывает значительный эффект тренировочных дней ( F (9 162) = 102,17, p <0,0001). Поскольку начальное значение ошибки различается для четырех групп, неудивительно, что также наблюдался значительный групповой эффект ( F (3,198) = 14.
A показывает, что действительно доля испытаний, в которых выполнялась УЗИ, уменьшилась по сравнению с тренировкой, что указывает на увеличение количества испытаний, в которых целевая амплитуда была достигнута или превышена. Для этого показателя наблюдалось значительное влияние групп ( F (3,198) = 7,13, p = 0,0047) и тренировочного дня ( F (9,162) = 70,93, p <0,0001 ), но сравнение взаимодействий не было статистически надежным ( F (27,162) = 1.67, p = 0,0545). Во-вторых, должно происходить уменьшение средней ошибки, то есть среднего абсолютного значения разницы между амплитудой отклика и целевой амплитудой. Этот показатель отображается как функция количества тренировочных дней в B . На общее уменьшение средней ошибки указывает значительный эффект тренировочных дней ( F (9 162) = 102,17, p <0,0001). Поскольку начальное значение ошибки различается для четырех групп, неудивительно, что также наблюдался значительный групповой эффект ( F (3,198) = 14. 41, p <0,0001) и значительный эффект группы × тренировочные дни ( F (27,162) = 13,6, p <0,0001).
41, p <0,0001) и значительный эффект группы × тренировочные дни ( F (27,162) = 13,6, p <0,0001).
A , B , Уменьшение вероятности доставки в США ( A ) и средней ошибки ( B ) в зависимости от 10 дней обучения для четыре группы амплитуд (2, 3, 4 и 5 мм). Снижение вероятности УЗИ ( A ) указывает на увеличение доли ответов, которые соответствуют целевой амплитуде или превышают ее.Все четыре группы показали значительное снижение вероятности поражения в США, достигнув значений от 21 до 44%. B , Средняя ошибка уменьшилась до аналогичных значений во всех четырех группах.
В целом, эти анализы показывают, что (1) группы, разработанные с разницей в тренировках по амплитуде и величине ответа, (2) доля испытаний, в которых проводилось УЗИ, и средняя ошибка уменьшались за время обучения, и (3) там Нет различий в вероятности ответа для трех групп. Поскольку наиболее простой способ оценить свойства этого обучения – это оценить производительность в стабильном или близком к нему состоянии, далее мы сосредоточили анализ на данных за последние 2 дня обучения.
Поскольку наиболее простой способ оценить свойства этого обучения – это оценить производительность в стабильном или близком к нему состоянии, далее мы сосредоточили анализ на данных за последние 2 дня обучения.
Анализ асимптотической производительности
Для анализа стабильной производительности данные были объединены с 9-го и 10-го дней сбора данных для каждого субъекта. Этот анализ выявил четкие различия в амплитуде и величине CR между четырьмя группами ( A , B ).Отдельные односторонние дисперсионные анализы для амплитуды ( F (3,18) = 19,53, p <0,0001) и величины ( F = 37,25, p <0,0001) подтверждают надежность этих различий. Для измерения величины все шесть парных сравнений (критерий Тьюки, p <0,05) были надежными, а для амплитуды только сравнение 3 и 4 мм не было надежным. В отличие от этих различий, не было достоверных различий в измерении средней ошибки между четырьмя группами ( C ). В качестве финального теста на различия в амплитуде и величине ответов по четырем целевым амплитудным группам были рассчитаны нормализованные совокупные частотные распределения для каждой меры ответа ( G ). Тест Колмогорова – Смирнова ( p <0,005) показал, что все шесть парных сравнений для каждой группы были статистически надежными.
В качестве финального теста на различия в амплитуде и величине ответов по четырем целевым амплитудным группам были рассчитаны нормализованные совокупные частотные распределения для каждой меры ответа ( G ). Тест Колмогорова – Смирнова ( p <0,005) показал, что все шесть парных сравнений для каждой группы были статистически надежными.
Сравнения четырех амплитудных групп при стабильных показателях, взятых для каждого животного с 9-го и 10-го дней. A , Установившаяся амплитуда ответа различалась для четырех групп.Четыре пунктирные линии указывают целевую амплитуду для каждой группы. Все попарные сравнения (критерий Тьюки, p <0,05) были значимыми, за исключением сравнения между группами 3 и 4 мм. B , тот же формат, что и A для данных величины, которые представляют собой среднюю амплитуду всех CR без исключения не-CR. Все попарные сравнения были значимыми (критерий Тьюки, p <0,05). C , Стационарная средняя ошибка (абсолютное значение) существенно не различалась для четырех групп. D , Наблюдалась общая тенденция увеличения доли испытаний, в которых использовались УЗИ, с увеличением амплитуды цели. Единственное парное сравнение, которое было статистически надежным, было 2 против 5 мм. E , Все четыре группы показывают сопоставимые характеристики с точки зрения вероятности CR; между четырьмя группами не было значительных различий. F , Была общая тенденция к увеличению пиковой скорости CR по мере увеличения целевой амплитуды.Единственное достоверное (критерий Тьюки, p <0,05) сравнение было 2 против 5 мм. G , Суммарная доля ответов как функция амплитуды условного ответа (вверху) и величины (внизу) для четырех групп. Каждый CR для каждого животного помещали в бункер (ширина бункера 0,2 мм). Все шесть парных сравнений являются статистически надежными (критерий Колмогорова – Смирнова, p <0,005) для амплитуды, а также для измерения величины (кроме 4 против 5 мм, p <0.
C , Стационарная средняя ошибка (абсолютное значение) существенно не различалась для четырех групп. D , Наблюдалась общая тенденция увеличения доли испытаний, в которых использовались УЗИ, с увеличением амплитуды цели. Единственное парное сравнение, которое было статистически надежным, было 2 против 5 мм. E , Все четыре группы показывают сопоставимые характеристики с точки зрения вероятности CR; между четырьмя группами не было значительных различий. F , Была общая тенденция к увеличению пиковой скорости CR по мере увеличения целевой амплитуды.Единственное достоверное (критерий Тьюки, p <0,05) сравнение было 2 против 5 мм. G , Суммарная доля ответов как функция амплитуды условного ответа (вверху) и величины (внизу) для четырех групп. Каждый CR для каждого животного помещали в бункер (ширина бункера 0,2 мм). Все шесть парных сравнений являются статистически надежными (критерий Колмогорова – Смирнова, p <0,005) для амплитуды, а также для измерения величины (кроме 4 против 5 мм, p <0. 05).
05).
A – C в совокупности предполагают, что, хотя средняя ошибка амплитуды была сопоставима в четырех группах, источники ошибок были разными. Ответы для групп 2 и 3 мм имели тенденцию выходить за цель, тогда как ответы для группы 5 мм имели тенденцию выходить за пределы цели. Эту тенденцию можно вывести из сравнения вероятности в США, показанного в D , которое отражает долю ответов, которые не достигли целевого значения. Несмотря на четкую тенденцию к увеличению этого показателя с целевой амплитудой, различия между группами только приближались к значимости ( F (3,18) = 3.44, p = 0,08). Только группа 2 мм показала значительную тенденцию к превышению целевого значения, потому что отдельные тесты t для каждой группы выявили только достоверное отличие от среднего значения популяции в 50% (равные реакции недостижения и превышения) для группы 2 мм ( t ). (5) = 3,47, p <0,05). Несмотря на разницу в амплитуде и величине ответа, группы показали сопоставимые показатели ( F (3,18) = 1,799, p = 0. 37) с точки зрения вероятности CR ( E ). Наконец, F показывает, что существует также общая тенденция увеличения максимальной скорости КЛ с целевой амплитудой ( F (3,18) = 7,81, p = 0,003). Парные сравнения показали, что различия 2 по сравнению с 4 мм и 2 по сравнению с 5 мм были надежными (критерий Тьюки, p <0,05).
37) с точки зрения вероятности CR ( E ). Наконец, F показывает, что существует также общая тенденция увеличения максимальной скорости КЛ с целевой амплитудой ( F (3,18) = 7,81, p = 0,003). Парные сравнения показали, что различия 2 по сравнению с 4 мм и 2 по сравнению с 5 мм были надежными (критерий Тьюки, p <0,05).
Предыдущие исследования показали, что точное время CR является характерной чертой кондиционирования век (Smith, 1968; Smith et al., 1969; Милленсон и др., 1977; Маук и Руис, 1992). Время ожидания и время нарастания ответов систематически меняются в зависимости от межстимульного интервала, так что ответы имеют тенденцию достигать пика около времени доставки УЗИ. Мы исследовали различные меры времени ответа для четырех целевых амплитудных групп и не обнаружили достоверных различий во времени ответа ( A ). Ни время ожидания начала, ни время ожидания критерия, время ожидания максимальной скорости, ни время ожидания ответа пика не показали статистически достоверных различий между группами. Когда ответы были разделены в зависимости от того, превышают ли они цель (испытания в США) или ниже цели (испытания в США), наблюдались значительные различия во времени ответов во всех четырех группах ( B ). Отдельные двухсторонние смешанные ANOVA для задержки начала и задержки до максимальной скорости показали только значимое влияние типа ответа (не в США по сравнению с США; начало, F (1,18) = 100,6, p < 0,0001; пиковая скорость, F = 49.04, p <0,0001). Этот анализ показывает, что в целом ответы, которые соответствовали целевой амплитуде или превышали ее, имели более короткие задержки, чем ответы, которые не соответствовали целевой амплитуде.
Когда ответы были разделены в зависимости от того, превышают ли они цель (испытания в США) или ниже цели (испытания в США), наблюдались значительные различия во времени ответов во всех четырех группах ( B ). Отдельные двухсторонние смешанные ANOVA для задержки начала и задержки до максимальной скорости показали только значимое влияние типа ответа (не в США по сравнению с США; начало, F (1,18) = 100,6, p < 0,0001; пиковая скорость, F = 49.04, p <0,0001). Этот анализ показывает, что в целом ответы, которые соответствовали целевой амплитуде или превышали ее, имели более короткие задержки, чем ответы, которые не соответствовали целевой амплитуде.
Меры времени отклика по сравнению с четырьмя группами амплитуды. A , Четыре графика показывают задержку до начала, задержку до критерия, задержку до пиковой скорости и задержку до пиковой амплитуды (для испытаний за пределами США). Статистически достоверной разницы между группами по этим показателям ответа не было. B , Разница во времени испытаний в США и за пределами США. Раздельный анализ исследований в УЗИ (амплитуда ниже целевого) и неамериканских исследований (амплитуда на уровне или выше) выявил общую тенденцию к более быстрым и коротким задержкам ответов в неамериканских исследованиях. Все восемь сравнений (4 группы × 2 меры ответа) были статистически надежными (парный тест t , p <0,01).
B , Разница во времени испытаний в США и за пределами США. Раздельный анализ исследований в УЗИ (амплитуда ниже целевого) и неамериканских исследований (амплитуда на уровне или выше) выявил общую тенденцию к более быстрым и коротким задержкам ответов в неамериканских исследованиях. Все восемь сравнений (4 группы × 2 меры ответа) были статистически надежными (парный тест t , p <0,01).
Примеры необработанных ответов от одного испытуемого из каждой амплитудной группы (день 10 в каждом случае) показаны в.Для каждого субъекта ответы, которые соответствовали целевой амплитуде или превышали ее, показаны в левом столбце, а те, которые не соответствовали целевой амплитуде (и, следовательно, были доставлены УЗИ), показаны в правом столбце. Эти данные показывают общую тенденцию реакции к перерегулированию для меньшей амплитуды цели и к недолету для цели 5 мм. Вместе эти анализы демонстрируют, что процедура тренировки целевой амплитуды может использоваться для получения разумного экспериментального контроля над амплитудой ответа при кондиционировании век, что средняя ошибка амплитуды сопоставима для целей от 2 до 5 мм, и что эти различия возникают на сопоставимых уровнях CR. и сопоставимые статистические данные о сроках проведения CR.
и сопоставимые статистические данные о сроках проведения CR.
Пример необработанных ответов от четырех животных, по одному от каждой группы амплитуд, взятых с 10-го дня обучения. Каждая развертка представляет собой отдельную реакцию века (первая проба впереди), причем черная область каждой развертки указывает продолжительность CS. Ответы были разделены на испытания, проводимые за пределами США (левый столбец) и испытания в США (правый столбец). Последовательность испытаний каждого животного в США и за пределами США очевидна по интервалу между испытаниями. Обратите внимание на общую тенденцию к большему количеству испытаний без УЗИ с малыми амплитудами цели и к большему количеству испытаний УЗИ с большими амплитудами цели.Компонент рефлекторного ответа каждого испытания виден в светло-серой области испытаний «With-US».
Амплитуда отклика после повторного обнаружения новой цели
Практическая и концептуальная важность тренировки амплитуды целевой будет уменьшена, если амплитуды отклика цели не могут быть получены и повторно получены в соответствии с планом эксперимента. Поэтому мы тестировали каждое животное, используя новую целевую амплитуду, отличную от начальной. После 10 дней обучения до начальной целевой амплитуды каждый субъект был обучен в течение 10 дополнительных дней с использованием новой целевой амплитуды.Новые целевые амплитуды для групп 2, 3, 4 и 5 мм, соответственно, составляли 4, 5, 2 и 3 мм. показывает сравнение амплитуд ответа для этих четырех групп для начальной и новой целевой амплитуд. Как и выше, эти данные включали объединенное среднее значение 9 и 10 дней для каждого субъекта. Мы начали с того, что спросили, есть ли различия между получением и повторным обнаружением для каждой из четырех целевых амплитуд (левый график). Это сравнение между субъектами сравнивает каждую цель, когда она использовалась при захвате, по сравнению с повторным захватом (т.е., получение от группы 2 мм и повторное обнаружение на цели 2 мм, первоначально группы 4 мм). Двусторонний дисперсионный анализ выявил только основной эффект целевой амплитуды ( F (3,36) = 37,06, p <0,0001), эффекты захвата по сравнению с повторным захватом и взаимодействия не были значительными, и не было надежных различия в амплитудах, полученных во время захвата, по сравнению с повторным обнаружением новой цели.
Поэтому мы тестировали каждое животное, используя новую целевую амплитуду, отличную от начальной. После 10 дней обучения до начальной целевой амплитуды каждый субъект был обучен в течение 10 дополнительных дней с использованием новой целевой амплитуды.Новые целевые амплитуды для групп 2, 3, 4 и 5 мм, соответственно, составляли 4, 5, 2 и 3 мм. показывает сравнение амплитуд ответа для этих четырех групп для начальной и новой целевой амплитуд. Как и выше, эти данные включали объединенное среднее значение 9 и 10 дней для каждого субъекта. Мы начали с того, что спросили, есть ли различия между получением и повторным обнаружением для каждой из четырех целевых амплитуд (левый график). Это сравнение между субъектами сравнивает каждую цель, когда она использовалась при захвате, по сравнению с повторным захватом (т.е., получение от группы 2 мм и повторное обнаружение на цели 2 мм, первоначально группы 4 мм). Двусторонний дисперсионный анализ выявил только основной эффект целевой амплитуды ( F (3,36) = 37,06, p <0,0001), эффекты захвата по сравнению с повторным захватом и взаимодействия не были значительными, и не было надежных различия в амплитудах, полученных во время захвата, по сравнению с повторным обнаружением новой цели. Правая часть показывает те же данные, построенные заново, чтобы выделить сравнения амплитуд, полученных для сбора и повторного сбора данных в одних и тех же группах.Четыре отдельных парных теста t ( p <0,01) показали, что для каждой группы амплитуда ответов, полученных при обнаружении одной цели, отличалась от тех, которые были получены при повторном обнаружении на новую цель. Эти анализы показывают, что как получение первой целевой амплитуды, так и повторное обнаружение новой целевой амплитуды вызвали значительную разницу в амплитуде ответа для каждой целевой амплитудной группы и что не было никаких различий в этих тенденциях между захватом и повторным захватом.
Правая часть показывает те же данные, построенные заново, чтобы выделить сравнения амплитуд, полученных для сбора и повторного сбора данных в одних и тех же группах.Четыре отдельных парных теста t ( p <0,01) показали, что для каждой группы амплитуда ответов, полученных при обнаружении одной цели, отличалась от тех, которые были получены при повторном обнаружении на новую цель. Эти анализы показывают, что как получение первой целевой амплитуды, так и повторное обнаружение новой целевой амплитуды вызвали значительную разницу в амплитуде ответа для каждой целевой амплитудной группы и что не было никаких различий в этих тенденциях между захватом и повторным захватом.
Сравнение устойчивых амплитуд ответа после первоначального сбора данных (темно-серый; те же данные, что и A ) и у тех же животных после 10 дней повторного обнаружения с новой целевой амплитудой. Начальная амплитуда цели и амплитуда цели повторного обнаружения были, соответственно, для четырех групп (от 2 до 4 мм, от 3 до 5 мм, от 4 до 2 мм и от 5 до 3 мм). Как и в случае, данные получены из среднего значения для каждого животного за 9-й и 10-й дни захвата (и повторного захвата). Слева: сравнение идентичных целевых амплитуд для захвата и повторного захвата.Статистически достоверных различий для каждой целевой амплитуды между данными сбора и повторного обнаружения не было ( t тестов). Целевая амплитуда обозначена пунктирными линиями. Справа: сравнение данных приобретения и повторного приобретения для одних и тех же групп животных. Различные целевые амплитуды захвата и повторного захвата для каждой группы снова обозначены пунктирными линиями. Парные тесты t ( p <0,01) показали статистически достоверные различия между амплитудами сбора и повторного обнаружения для каждой группы.
Как и в случае, данные получены из среднего значения для каждого животного за 9-й и 10-й дни захвата (и повторного захвата). Слева: сравнение идентичных целевых амплитуд для захвата и повторного захвата.Статистически достоверных различий для каждой целевой амплитуды между данными сбора и повторного обнаружения не было ( t тестов). Целевая амплитуда обозначена пунктирными линиями. Справа: сравнение данных приобретения и повторного приобретения для одних и тех же групп животных. Различные целевые амплитуды захвата и повторного захвата для каждой группы снова обозначены пунктирными линиями. Парные тесты t ( p <0,01) показали статистически достоверные различия между амплитудами сбора и повторного обнаружения для каждой группы.
Стимуляция мшистых волокон в качестве CS
Значительные доказательства указывают на то, что приобретение и экспрессия CR во время формирования павлового (отложенного) века опосредуется пластичностью мозжечка. Поскольку обусловленность целевой амплитуды, строго говоря, не является павловской, мы провели два дополнительных эксперимента, чтобы проверить возможность того, что обучение в этих условиях опосредуется нецеребеллярными механизмами. Способность заменять стимуляцию мшистых волокон тонусом CS – одно из наиболее убедительных доказательств, подтверждающих, что мозжечковые механизмы являются основой кондиционирования век.Таким образом, мы сравнили эффекты тренировки с целевой амплитудой 3 мм, используя тон CS в одной группе и стимуляцию мшистых волокон в качестве CS во второй группе. В остальном процедуры были идентичны процедурам, использованным выше для группы 3 мм. показывает результаты за 10 дней тренировок для этих двух групп. A – D сравнивает эти две группы за 10 дней обучения, а E показывает сравнения в установившемся состоянии, как в предыдущем эксперименте. Отдельные двухфакторные смешанные ANOVA для четырех мер реакции показали только основные эффекты тренировочных дней (амплитуда, F (9,54) = 30.54; ошибка, F = 17,71; Вероятность CR, F = 29,09; Вероятность США – 16,62; p <0,0001 во всех случаях). Ни один из основных эффектов группы или эффектов взаимодействия не был значимым, равно как и сравнение тона и мшистого волокна ( E ) для восьми показателей реакции, которые были проанализированы для данных устойчивого состояния (дни 9 и 10, t тесты, p > 0,05).
Способность заменять стимуляцию мшистых волокон тонусом CS – одно из наиболее убедительных доказательств, подтверждающих, что мозжечковые механизмы являются основой кондиционирования век.Таким образом, мы сравнили эффекты тренировки с целевой амплитудой 3 мм, используя тон CS в одной группе и стимуляцию мшистых волокон в качестве CS во второй группе. В остальном процедуры были идентичны процедурам, использованным выше для группы 3 мм. показывает результаты за 10 дней тренировок для этих двух групп. A – D сравнивает эти две группы за 10 дней обучения, а E показывает сравнения в установившемся состоянии, как в предыдущем эксперименте. Отдельные двухфакторные смешанные ANOVA для четырех мер реакции показали только основные эффекты тренировочных дней (амплитуда, F (9,54) = 30.54; ошибка, F = 17,71; Вероятность CR, F = 29,09; Вероятность США – 16,62; p <0,0001 во всех случаях). Ни один из основных эффектов группы или эффектов взаимодействия не был значимым, равно как и сравнение тона и мшистого волокна ( E ) для восьми показателей реакции, которые были проанализированы для данных устойчивого состояния (дни 9 и 10, t тесты, p > 0,05). Эти данные исключают факторы, расположенные выше мозжечка, как способствующие приобретению амплитуды ответа, и подтверждают гипотезу о том, что кондиционирование век к целевой амплитуде опосредуется мозжечком.
Эти данные исключают факторы, расположенные выше мозжечка, как способствующие приобретению амплитуды ответа, и подтверждают гипотезу о том, что кондиционирование век к целевой амплитуде опосредуется мозжечком.
Получение целевой амплитуды (3 мм) для животных, обученных с помощью тонального CS и стимуляции мшистыми волокнами в качестве CS. A , Амплитуда CR за 10 дней обучения. B , Средняя ошибка (абсолютное значение) за 10 дней обучения. MF, Мшистое волокно. C , D , вероятность CR ( C ) и вероятность США ( D ) за 10 дней обучения. E , Сравнение между группами CS тональной стимуляции и CS со стимуляцией мшистыми волокнами для различных мер реакции, взятых с 9-го и 10-го дней тренировки.Ни одно из шести сравнений групп не было статистически надежным (тест t , p > 0,05).
Обратимая инактивация мозжечка
В качестве дополнительной проверки гипотезы о том, что мозжечок необходим для кондиционирования до заданной амплитуды, экспрессия CR была протестирована на четырех животных, обученных тону, и четырех животных, обученных мшистым волокнам, до и после инактивации. мозжечкового выхода через инфузию мусцимола, агониста ГАМК, в переднее межположительное ядро ().Субъекты в обеих группах тестировались в отдельные дни с инфузиями носителя или инфузиями мусцимола (порядок уравновешивался для каждой группы). График средних показателей по группе (в центре) показывает, что для обеих групп не было достоверных изменений после инфузии носителя, тогда как инфузии мусцимола отменяли экспрессию CR. Необработанные данные для одного субъекта стимуляции мшистых волокон для инфузий носителя (слева) и мусцимола (справа) показаны вверху. Пример канюли (слева) и стимулирующих электродов от того же пациента показаны внизу.
Необходимость мозжечка для выражения приобретенных ответов на заданную амплитуду 3 мм. Две группы, одна из которых использовала тональный CS, а другая, в которой стимуляция волокон использовалась в качестве CS, были протестированы в отдельные дни либо с инфузией носителя, либо с инфузией мусцимола, агониста ГАМК. На двух графиках показаны средние показатели по группе отсутствия эффекта от инфузии носителя (слева) и отмены CR с помощью инфузии мусцимола (справа). Время настаивания в каждом случае указано черной стрелкой.Примеры исходных данных от одного животного показаны выше: следы слева – это инфузия носителя, а следы справа – инфузия мусцимола. В каждом случае ответы до инфузии находятся впереди (первое испытание впереди), а реакции после инфузии – сзади. На двух нижних фотографиях показано размещение инфузионной канюли (слева) и стимулирующих электродов из мшистых волокон (справа) для животного, исходные данные которого показаны выше. Этому животному были имплантированы два электрода из покрытых мхом волокон; только один был использован для этого эксперимента.MF, Мшистые волокна.
Две группы, одна из которых использовала тональный CS, а другая, в которой стимуляция волокон использовалась в качестве CS, были протестированы в отдельные дни либо с инфузией носителя, либо с инфузией мусцимола, агониста ГАМК. На двух графиках показаны средние показатели по группе отсутствия эффекта от инфузии носителя (слева) и отмены CR с помощью инфузии мусцимола (справа). Время настаивания в каждом случае указано черной стрелкой.Примеры исходных данных от одного животного показаны выше: следы слева – это инфузия носителя, а следы справа – инфузия мусцимола. В каждом случае ответы до инфузии находятся впереди (первое испытание впереди), а реакции после инфузии – сзади. На двух нижних фотографиях показано размещение инфузионной канюли (слева) и стимулирующих электродов из мшистых волокон (справа) для животного, исходные данные которого показаны выше. Этому животному были имплантированы два электрода из покрытых мхом волокон; только один был использован для этого эксперимента.MF, Мшистые волокна.
В совокупности исследования стимуляции мшистых волокон и инактивации глубоких ядер показывают (1), что мозжечок необходим для экспрессии CR, продуцируемых кондиционированием века до целевой амплитуды (результаты инфузии), и (2) что механизмы, ответственные за приобретение и экспрессия целевой амплитуды ответа не находятся в пути CR выше мозжечка (результаты стимуляции мшистыми волокнами).
Обсуждение
Мы адаптировали традиционные процедуры кондиционирования павловских век, чтобы ввод ошибки, определяемый США, зависел от амплитуды CR.Пропустив УЗИ в исследованиях, в которых CR соответствует или превышает заданную целевую амплитуду (и представив УЗИ в противном случае), мы показали, что результирующие условные реакции век имеют тенденцию к целевой амплитуде. Это верно как для получения исходной целевой амплитуды, так и для повторного обнаружения с использованием другой цели. Несмотря на то, что наблюдалась тенденция к перерегулированию при малых амплитудах цели и к недолету с наибольшей целью, средняя абсолютная ошибка не различалась для групп, обученных для целевых амплитуд от 2 до 5 мм. Эти данные показывают, что, адаптируя процедуры кондиционирования век к естественным условиям обучения мозжечка, амплитуду CR можно поставить под экспериментальный контроль. Способность устранять эти CR с помощью инактивации мозжечка и способность заменять стимуляцию мшистых волокон тональным CS подтверждают гипотезу о том, что, как и в случае кондиционирования века в форме павлова, кондиционирование века до целевой амплитуды опосредуется мозжечком. Эти результаты формируют основу для систематических и контролируемых исследований того, как мозжечок обучается, сохраняет и выражает надлежащую амплитуду откликов, которые он адаптирует.
Эти данные показывают, что, адаптируя процедуры кондиционирования век к естественным условиям обучения мозжечка, амплитуду CR можно поставить под экспериментальный контроль. Способность устранять эти CR с помощью инактивации мозжечка и способность заменять стимуляцию мшистых волокон тональным CS подтверждают гипотезу о том, что, как и в случае кондиционирования века в форме павлова, кондиционирование века до целевой амплитуды опосредуется мозжечком. Эти результаты формируют основу для систематических и контролируемых исследований того, как мозжечок обучается, сохраняет и выражает надлежащую амплитуду откликов, которые он адаптирует.
Значение для кондиционирования век и мозжечка
Необходимость мозжечка для кондиционирования века с задержкой павлова подтверждается большим количеством данных. Настоящие результаты показывают, что непредвиденные обстоятельства Павлова, при которых доставка УЗИ не зависит от CR, не являются особенностью, которая делает мозжечок необходимым. Поскольку, строго говоря, процедуры тренировки до целевой амплитуды включают в себя оперантные непредвиденные обстоятельства, необходимость мозжечка в кондиционировании век связана с природой стимулов и системой реакции, а не с природой непредвиденных обстоятельств, которые управляют представлением усиливающий стимул.
Настоящие данные также обращаются к распространенному заблуждению о кондиционировании век, а именно, что это рефлекторное кондиционирование или кондиционирование моргания глаз. Павлов выступил с идеей, что классическое кондиционирование включает в себя пути, активируемые CS, обретающие способность активировать путь, который опосредует рефлекторный ответ (Павлов, 1927). Нервные пути, которые, как известно, опосредуют экспрессию CR и рефлекторные мигания, вызываемые США, не согласуются с гипотезой Павлова. Рефлекторные мигания, вызываемые первичными афферентами, проходящими через верхний глазничный нерв, опосредуются двумя путями (Pellegrini et al., 1995). Первичные афференты проецируются как на каудально-межполярную пограничную область спинномозгового ядра тройничного нерва, так и на дорсальный рог, сегмент С1 спинного мозга. Обе эти области проецируются в область orbicularis oculi лицевого ядра. Эксперименты с поражением показали, что спинальный тройничный путь опосредует начальный компонент R1 рефлекторных морганий, а путь спинного мозга опосредует более длинный латентный компонент R2 (Pelligrini et al. , 1995). Путь CR, выступающий из переднего межположительного ядра через красное ядро, по-видимому, соединяется с этим путем в двигательных или премоторных нейронах лицевого ядра.Следовательно, рефлекторные мигания и CRs опосредуются совершенно разными нервными путями и, как можно ожидать, будут проявлять совершенно разные свойства, как было четко установлено в исследованиях на людях (Schade Powers et al., 2010). Поэтому условные реакции век более правильно охарактеризовать как «реакции век», чем «моргания глаз», и настоящие данные демонстрируют, что амплитуда этих ответов век, как и большинство других реакций скелетных мышц, может быть изучена и опосредована обучением мозжечка.
, 1995). Путь CR, выступающий из переднего межположительного ядра через красное ядро, по-видимому, соединяется с этим путем в двигательных или премоторных нейронах лицевого ядра.Следовательно, рефлекторные мигания и CRs опосредуются совершенно разными нервными путями и, как можно ожидать, будут проявлять совершенно разные свойства, как было четко установлено в исследованиях на людях (Schade Powers et al., 2010). Поэтому условные реакции век более правильно охарактеризовать как «реакции век», чем «моргания глаз», и настоящие данные демонстрируют, что амплитуда этих ответов век, как и большинство других реакций скелетных мышц, может быть изучена и опосредована обучением мозжечка.
Значение для обучения мозжечка усилению или амплитуде
Вопрос о том, как и где мозжечок обучается и сохраняет усиление или амплитуду ответа, взаимосвязан с вопросом о месте (ах) пластичности, ответственных за обучение мозжечка. На этот вопрос лучше всего ответить с помощью исследований, включающих анализ адаптации VOR (Raymond et al. , 1996; Boyden et al., 2004). Первоначальные усилия в этой области вызвали серьезные дискуссии о том, происходит ли пластичность в коре мозжечка или в вестибулярном ядре, чьи связи и схема параллельны глубоким ядрам мозжечка (Lisberger and Sejnowski, 1992, 1993; Ito, 1993).Более поздние теории предполагают участие пластичности как в коре мозжечка, так и в глубоком ядре (Perrett et al., 1993; Raymond et al., 1996; Mauk and Donegan, 1997). Когда задействовано более одного сайта пластичности, становится важным вопрос об относительном вкладе этих двух сайтов. Относительный вклад коры мозжечка и механизмов пластичности глубокого ядра мозжечка в усиление или амплитуду ответа остается нерешенным, и есть указания на то, что относительный вклад может варьироваться сложным образом в зависимости от обстоятельств (Mauk, 1997).Кондиционирование век до целевой амплитуды обеспечивает способ решения этих вопросов с помощью относительно простой и экспериментально управляемой системы, которая поддается параметрическому поведенческому анализу, обратимым повреждениям, in vivo записи и даже крупномасштабному компьютерному моделированию.
, 1996; Boyden et al., 2004). Первоначальные усилия в этой области вызвали серьезные дискуссии о том, происходит ли пластичность в коре мозжечка или в вестибулярном ядре, чьи связи и схема параллельны глубоким ядрам мозжечка (Lisberger and Sejnowski, 1992, 1993; Ito, 1993).Более поздние теории предполагают участие пластичности как в коре мозжечка, так и в глубоком ядре (Perrett et al., 1993; Raymond et al., 1996; Mauk and Donegan, 1997). Когда задействовано более одного сайта пластичности, становится важным вопрос об относительном вкладе этих двух сайтов. Относительный вклад коры мозжечка и механизмов пластичности глубокого ядра мозжечка в усиление или амплитуду ответа остается нерешенным, и есть указания на то, что относительный вклад может варьироваться сложным образом в зависимости от обстоятельств (Mauk, 1997).Кондиционирование век до целевой амплитуды обеспечивает способ решения этих вопросов с помощью относительно простой и экспериментально управляемой системы, которая поддается параметрическому поведенческому анализу, обратимым повреждениям, in vivo записи и даже крупномасштабному компьютерному моделированию.
Нарушение классического кондиционирования век после поражения мозжечка: повреждение системы следа памяти или простой дефицит работоспособности?
За последние 10 лет ряд лабораторий сообщил, что классически обусловленные реакции скелетных мышц, такие как условные реакции мигательной мембраны / века, критически зависят от активности мозжечка.Например, было показано, что односторонние поражения межположительного ядра мозжечка предотвращают приобретение и отменяют сохранение условной реакции века на стороне, ипсилатеральной по отношению к поражениям, не влияя на условную реакцию (CR) на противоположной стороне. Кроме того, записи исследований с участием межположительного ядра постоянно выявляют паттерны нейрональной разгрузки, которые предсказывают выполнение CR. Исследования повреждений и записи обычно приводятся как доказательство того, что пластичность мозжечка критически важна для обучения и запоминания классически обусловленных реакций. Эта интерпретация была недавно оспорена Уэлшем и Харви (1989a), которые утверждали, что поражения мозжечка просто вызывают снижение производительности, и предположили, что роль мозжечка заключается не в процессах обучения и памяти, связанных с CR, а только в выполнении моргания глазом. отклик. Здесь представлены три эксперимента, которые предоставляют дополнительные убедительные доказательства критической роли мозжечка в обучении и памяти Павловской CR. Эти эксперименты включают (1) демонстрацию полной и постоянной отмены CR после соответствующих межположительных поражений, (2) неспособность найти систематические или стойкие снижения амплитуды безусловного ответа (т.е., рефлекс моргания глаза) после соответствующего межположительного поражения, и (3) наблюдения дифференциальных эффектов на CR и безусловный ответ после того, как поражения были помещены в популяции мотонейронов, ответственных за выполнение реакции моргания глаз. Эти данные обсуждаются в контексте вопросов производительности и обучения; Данные, представленные здесь, исключают возможность того, что устранение межположительного поражения CR моргания глаз просто связано с влиянием поражения на производительность.
Эта интерпретация была недавно оспорена Уэлшем и Харви (1989a), которые утверждали, что поражения мозжечка просто вызывают снижение производительности, и предположили, что роль мозжечка заключается не в процессах обучения и памяти, связанных с CR, а только в выполнении моргания глазом. отклик. Здесь представлены три эксперимента, которые предоставляют дополнительные убедительные доказательства критической роли мозжечка в обучении и памяти Павловской CR. Эти эксперименты включают (1) демонстрацию полной и постоянной отмены CR после соответствующих межположительных поражений, (2) неспособность найти систематические или стойкие снижения амплитуды безусловного ответа (т.е., рефлекс моргания глаза) после соответствующего межположительного поражения, и (3) наблюдения дифференциальных эффектов на CR и безусловный ответ после того, как поражения были помещены в популяции мотонейронов, ответственных за выполнение реакции моргания глаз. Эти данные обсуждаются в контексте вопросов производительности и обучения; Данные, представленные здесь, исключают возможность того, что устранение межположительного поражения CR моргания глаз просто связано с влиянием поражения на производительность.
Обработка мозжечка, общая для задержки и отслеживания кондиционирования век
Введение
Павловское кондиционирование век оказалось полезным для анализа нейронных механизмов обучения и обработки информации в таких областях мозга, как гиппокамп, мозжечок и префронтальная кора.Задержка и отслеживание кондиционирования век различаются только относительной синхронизацией стимулов: для кондиционирования задержки условный стимул (CS) и безусловный стимул (US) перекрываются во времени, тогда как кондиционирование следа включает временной промежуток между смещением CS и началом US. Тем не менее, существуют важные различия в вовлечении структур переднего мозга (Christian and Thompson, 2003). Хотя мозжечок необходим для обоих, поражения в различных областях переднего мозга влияют на следы, но не задерживают кондиционирование (Solomon and Moore, 1975; Berger and Orr, 1983; Woodruff-Pak et al., 1985; Соломон и др., 1986; Мойер и др. , 1990; Kalmbach et al., 2009, 2010a).
, 1990; Kalmbach et al., 2009, 2010a).
Недавние исследования дают прямое механистическое объяснение зависимости переднего мозга от кондиционирования век. Используя стимуляцию мшистых волокон в качестве CS, Kalmbach et al. (2010b) установили, что для обучения мозжечка необходимо, чтобы входы мшистых волокон, активируемые CS, перекрывались во времени с входами качающихся волокон, активируемыми США. Впоследствии Kalmbach et al. (2011) показали, что два входа мшистых волокон, один для продолжительности CS и один, сохраняющийся в интервале следа, были достаточны для мозжечка, чтобы получить ответы со свойствами, которые количественно соответствуют ответам следа век с традиционным тональным CS.В совокупности эти находки подтверждают, что необходимый вклад переднего мозга заключается в обеспечении мозжечка входящими в состав мшистыми волокнами, которые сохраняются в интервале следов, чтобы задействовать обучение мозжечка (Clark and Squire, 1998; Siegel et al., 2012). Следовательно, с точки зрения мозжечка, этот постоянный вход трансформирует условное кондиционирование следа в условное кондиционирование задержки, так что общие мозжечковые механизмы будут работать в обеих парадигмах (Kalmbach et al. , 2010b, 2011; Siegel et al., 2012). Эта точка зрения подтверждается исследованиями, показывающими, что инактивация коры мозжечка вызывает параллельные эффекты во время задержки и отслеживания кондиционирования век на экспрессию и время условных ответов (CR) (Kalmbach et al., 2010а).
, 2010b, 2011; Siegel et al., 2012). Эта точка зрения подтверждается исследованиями, показывающими, что инактивация коры мозжечка вызывает параллельные эффекты во время задержки и отслеживания кондиционирования век на экспрессию и время условных ответов (CR) (Kalmbach et al., 2010а).
Это объяснение оспаривается исследованиями, показывающими, что поражения коры мозжечка не влияют на получение или своевременное проявление следовых реакций век, а также наблюдения, что различные части коры мозжечка могут опосредовать след или задержку (Woodruff-Pak et al., 1985, 2006; Кишимото и др., 2001а, б, в; Браун и др., 2010). Напротив, поражения коры мозжечка препятствуют приобретению и влияют на время уже усвоенных откликов задержки (Perrett et al., 1993; Perrett and Mauk, 1995; Garcia and Mauk, 1998; Garcia et al., 1999). Эти результаты были интерпретированы как доказательство того, что вклад и механизмы коры мозжечка различаются для задержки и отслеживания кондиционирования век (Takehara-Nishiuchi, 2018). Однако эта точка зрения, по-видимому, противоречит организации парасагиттальной зоны коры мозжечка, где зоны контролируют специфическую систему ответа (Eccles et al., 1967; Ekerot and Larson, 1979; Groenewegen et al., 1979). Если эта точка зрения верна, она также требует новых взглядов на обучение мозжечка в различных обстоятельствах.
Однако эта точка зрения, по-видимому, противоречит организации парасагиттальной зоны коры мозжечка, где зоны контролируют специфическую систему ответа (Eccles et al., 1967; Ekerot and Larson, 1979; Groenewegen et al., 1979). Если эта точка зрения верна, она также требует новых взглядов на обучение мозжечка в различных обстоятельствах.
Клетки Пуркинье (PCs) обеспечивают продукцию коры мозжечка, и предыдущие исследования установили, что зависимое от обучения снижение активности PC век контролирует кинематику задержки CRs (Halverson et al., 2015a). Чтобы разрешить противоречивые взгляды на участие мозжечка в кондиционировании следов, мы записали активность ПК век во время задержки и кондиционирования следов. Мы обнаружили, что взаимосвязь между CR век и индивидуальной активностью ПК имеет общие свойства, связанные с задержкой и условием следа.Модель обратной скорости стрельбы показала, что соответствие между связанной с обучением активностью ПК и кинематическими свойствами CR век не различается для задержки и следового кондиционирования. Случаи, когда один и тот же ПК был записан во время перехода от кондиционирования задержки к трассировке, показали, что ПК относятся к поведенческим реакциям одинаково для задержки и согласования трассировки. Эти данные подтверждают гипотезу о том, что отслеживание и задержка кондиционирования век задействуют одни и те же ПК и что кора мозжечка работает по аналогичным правилам для задержки и отслеживания кондиционирования век.
Случаи, когда один и тот же ПК был записан во время перехода от кондиционирования задержки к трассировке, показали, что ПК относятся к поведенческим реакциям одинаково для задержки и согласования трассировки. Эти данные подтверждают гипотезу о том, что отслеживание и задержка кондиционирования век задействуют одни и те же ПК и что кора мозжечка работает по аналогичным правилам для задержки и отслеживания кондиционирования век.
Результаты
Расположение и идентификация ПК век
Мы зарегистрировали в общей сложности 493 единицы, которые были идентифицированы как ПК на основании их пиковой активности и наличия сложных спайков. Используя критерии, описанные в материалах и методах, 46 из этих единиц были классифицированы как ПК век (см. Также Halverson et al., 2015a). Эти ПК век были зарегистрированы на 20 из 40 тетродов, имплантированных трем кроликам. Расчетное местоположение каждой из 46 записей обозначено красными точками на Рисунке 1 A .Многие точки перекрываются из-за плотной кластеризации мест записи. Эти участки были сгруппированы рядом с первичной трещиной в более глубоких частях долек HVI и HV передней доли, часто в борозде, которая простирается наиболее каудально к глубоким ядрам.
Эти участки были сгруппированы рядом с первичной трещиной в более глубоких частях долек HVI и HV передней доли, часто в борозде, которая простирается наиболее каудально к глубоким ядрам.
Записи коры мозжечка и классификация ПК век. A , Сагиттальный разрез коры мозжечка, показывающий конечное расположение (красные точки) тетродов, которые регистрировали активность ПК век.Большинство компьютерных записей век были сделаны в непосредственной близости (240 мкм), что привело к перекрытию в указанных местах. B , Пример окна среза кластера, построенного как пик к пику, показывающего одиночный блок (зеленый), изолированный от многоэлементной записи (черный). C , Изоляция сложных пиков (красный) и простых пиков (зеленый) от исходного кластера, разрезанного в B с использованием параметра позднего пика. Для каждого кластера также показаны цветные сигналы на четырех каналах тетрода. D , Многоблочная непрерывная запись с одиночного провода тетрода, показывающая простые (зеленые) и сложные (красный) пики от ПК, идентифицированные методом кластерной резки в B и C . E , гистограмма перистимула (интервалы 10 мс) и растровый график сложных спайков из записи ПК века во время сеанса кондиционирования века. Растровый график располагается последовательно, внизу – первая проба.Эта запись была классифицирована как ПК века на основании идентифицированных сложных спайков, вызванных ультразвуковой стимуляцией века (стрелка). F , Многоблочная непрерывная запись с одиночного провода тетрода, показывающая простые (зеленые точки) и сложные (красные точки) спайки от изолированного ПК века во время испытания кондиционирования одиночного следа, когда кролик показал CR. Серая линия представляет тон CS 500 мс; темно-серая линия представляет 50 мс США.
D , Многоблочная непрерывная запись с одиночного провода тетрода, показывающая простые (зеленые) и сложные (красный) пики от ПК, идентифицированные методом кластерной резки в B и C . E , гистограмма перистимула (интервалы 10 мс) и растровый график сложных спайков из записи ПК века во время сеанса кондиционирования века. Растровый график располагается последовательно, внизу – первая проба.Эта запись была классифицирована как ПК века на основании идентифицированных сложных спайков, вызванных ультразвуковой стимуляцией века (стрелка). F , Многоблочная непрерывная запись с одиночного провода тетрода, показывающая простые (зеленые точки) и сложные (красные точки) спайки от изолированного ПК века во время испытания кондиционирования одиночного следа, когда кролик показал CR. Серая линия представляет тон CS 500 мс; темно-серая линия представляет 50 мс США.
Ответы ПК века во время кондиционирования задержки и следа
Если кора мозжечка работает аналогично для кондиционирования века задержки и следа, то можно ожидать, что взаимосвязь между ответами ПК и CR век в двух парадигмах также должна быть аналогичной.Несмотря на то, что предыдущие исследования характеризовали эту взаимосвязь для CR задержки (Halverson et al., 2015a), мы регистрировали ПК как во время задержки, так и во время кондиционирования следа по следующим причинам: (1) для проведения внутрисубъектного сравнения ответов ПК во время выражения задержки и отслеживание CR, (2) для исследования ответов ПК во время перехода от задержки к ответу на трассировку и, когда это возможно, (3) для внутрикомпьютерного сравнения задержки и согласования трассировки, когда одна ячейка удерживалась в течение достаточного количества сеансов.
Первоначально каждый кролик был обучен с использованием сеансов кондиционирования с задержкой с тоном CS 550 мс в паре с совпадающей 50 мс стимуляцией век US, что дало ISI 500 мс, которое для удобства мы будем называть задержкой 500. Обучение с задержкой 500 продолжали до тех пор, пока каждый кролик не достиг высокого уровня условного ответа (более 80% CR во время сеанса) и пока не встретился хотя бы один век PC (рис. 2 A , внизу). После кондиционирования с задержкой 500 и во время сеанса, в котором присутствовал хотя бы один ПК века, тренировка была переключена на настройку трассировки (трасса 500/500, указывающая на тон 500 мс и интервал трассировки 500 мс).С дополнительными сеансами кондиционирования следов была получена стойкая экспрессия следов CR век. Тренировочные сессии Trace 500/500 продолжали записывать ПК век во время устойчивой экспрессии трассовых ответов.
Обучение с задержкой 500 продолжали до тех пор, пока каждый кролик не достиг высокого уровня условного ответа (более 80% CR во время сеанса) и пока не встретился хотя бы один век PC (рис. 2 A , внизу). После кондиционирования с задержкой 500 и во время сеанса, в котором присутствовал хотя бы один ПК века, тренировка была переключена на настройку трассировки (трасса 500/500, указывающая на тон 500 мс и интервал трассировки 500 мс).С дополнительными сеансами кондиционирования следов была получена стойкая экспрессия следов CR век. Тренировочные сессии Trace 500/500 продолжали записывать ПК век во время устойчивой экспрессии трассовых ответов.
A , Примеры CR век и компьютерных записей век во время перехода от хорошо натренированного на Delay 500 (внизу) к переходному сеансу Trace 500/500 (посередине) и хорошо натренированного на Trace 500 / 500 (вверху). В нижнем примере показано снижение активности ПК, связанное с экспрессией хорошо обученных CR во время задержки 500. Сеанс перехода показывает снижение активности ПК и CR, связанных с гашением ISI задержки 500 в течение первой половины сеанса. В верхнем примере показано, как уменьшение активности ПК переходит в интервал трассировки, чтобы соответствовать выражению хорошо обученных CR во время трассировки 500/500. На гистограммах перистимула (интервалы 10 мс) продолжительность тона CS представлена серым прямоугольником, а начало стимуляции века US представлено темно-серой линией для этого и всех последующих рисунков.Для поведенческих разверток синий цвет представляет длительность тонального стимула, а красный – интервал следа. Поведенческие развертки согласованы между испытаниями с растровыми графиками в каждом примере. B , Мгновенная скорость стрельбы (нормализованная к базовой линии) примерного ПК во время сеанса перехода от A . Угасание снижения активности ПК, связанного с выражением ранее обученных CR Delay 500, происходит в первой половине сеанса (пурпурный график) во время периода CS (серый прямоугольник).
Сеанс перехода показывает снижение активности ПК и CR, связанных с гашением ISI задержки 500 в течение первой половины сеанса. В верхнем примере показано, как уменьшение активности ПК переходит в интервал трассировки, чтобы соответствовать выражению хорошо обученных CR во время трассировки 500/500. На гистограммах перистимула (интервалы 10 мс) продолжительность тона CS представлена серым прямоугольником, а начало стимуляции века US представлено темно-серой линией для этого и всех последующих рисунков.Для поведенческих разверток синий цвет представляет длительность тонального стимула, а красный – интервал следа. Поведенческие развертки согласованы между испытаниями с растровыми графиками в каждом примере. B , Мгновенная скорость стрельбы (нормализованная к базовой линии) примерного ПК во время сеанса перехода от A . Угасание снижения активности ПК, связанного с выражением ранее обученных CR Delay 500, происходит в первой половине сеанса (пурпурный график) во время периода CS (серый прямоугольник). Активность ПК возвращается к исходному уровню после того, как CR погаснут во второй половине сеанса (черная кривая). C , Средняя нормализованная скорость активации ПК века и кинематические переменные CR века во время задержки 500 (оранжевый), задержки 1000 (голубой) и трассировки 500/500 (коричневый). Для этого и всех последующих рисунков штриховка вокруг средних линий представляет 95% доверительные интервалы для каждой кинематической переменной и активности ПК века. Темно-серые полосы представляют длительность CS, а светло-серые полосы представляют интервал трассировки.
Активность ПК возвращается к исходному уровню после того, как CR погаснут во второй половине сеанса (черная кривая). C , Средняя нормализованная скорость активации ПК века и кинематические переменные CR века во время задержки 500 (оранжевый), задержки 1000 (голубой) и трассировки 500/500 (коричневый). Для этого и всех последующих рисунков штриховка вокруг средних линий представляет 95% доверительные интервалы для каждой кинематической переменной и активности ПК века. Темно-серые полосы представляют длительность CS, а светло-серые полосы представляют интервал трассировки.
У двух кроликов тренировку затем смещали обратно к кондиционированию с задержкой с использованием ISI 1000 мс (задержка 1000), и эти сеансы продолжались до тех пор, пока не были обнаружены ПК век. Задержка 1000 позволяла сравнивать CR века и активность ПК века, обученную одному и тому же общему интервалу (1000 мс) во время задержки и кондиционирования трассы. Затем, как и раньше, обучение было переключено обратно на Trace 500/500, теперь с использованием другой частоты тонального сигнала CS для дальнейшего различения двух парадигм обучения и изучения ответов одного ПК на разные входные данные.Затем этим кроликам было проведено еще несколько сеансов кондиционирования Trace 500/500 для записи достаточного количества ПК век для сравнения их ответов во время асимптотического выполнения CR Trace 500/500 и Delay 1000.
Взаимосвязь между ПК-активностью века и условными реакциями век
Во время сеанса перехода от Delay 500 к Trace 500/500 задержка условных ответов век и связанное с ними снижение ПК-активности были погашены к середине сеанса ( Инжир.2 A , средний растровый график). Рисунок 2 B показывает среднюю частоту срабатывания ПК века для первой половины переходного сеанса, где присутствуют CR, по сравнению со второй половиной того же сеанса, где погаснут CR Delay 500. Поскольку CR Trace 500/500 появлялись на более поздних сессиях, время CR и связанное с ними снижение активности ПК века сдвигалось позже, обычно начиная с интервала следа (Рис.2 A , вверху), что характерно для века. CR со следовым кондиционированием.
Затем мы использовали средние за сеанс скорости активации ПК века, чтобы провести групповые сравнения ответов ПК века для Delay 500, Trace 500/500 и Delay 1000. Средняя мгновенная скорость активации ПК века, нормализованная к базовому уровню до CS, вдоль с положением и скоростью CR век для каждой тренировочной парадигмы показаны на Рисунке 2 C . Наложенные средние значения указывают на различия в ответах ПК век между парадигмами, обученными в разные интервалы, точно отслеживают время CR век для Delay 500, Trace 500/500 и Delay 1000.Заштрихованные области вокруг средних линий на Рисунке 2 C представляют 95% доверительные интервалы для каждой меры. Эти результаты, полученные на основе средних значений ПК и кинематических данных века, в целом согласуются с идеей о том, что ПК век контролируют время и экспрессию CR век как во время задержки, так и во время кондиционирования следа. Последующие анализы, описанные ниже, были направлены на дальнейшее количественное определение этого.
Контроль начала CR посредством активности ПК век
CR веков демонстрируют изменчивость времени начала ответа от испытания к испытанию, которая, что неудивительно, увеличивается с увеличением ISI.Эта вариабельность вводит степень усреднения артефакта, который может затруднить оценку того, насколько точны отношения между испытаниями между ответами ПК века и экспрессией CR век. Эта взаимосвязь между испытаниями визуально проиллюстрирована на Рисунке 3, A, и B , где показан пример ПК века с растровым графиком и поведение с испытаниями, показанными в хронологическом порядке (Рис. 3 A ) и снова. отсортированы по латентности начала CR (рис. 3 B ). Красные точки в каждой строке растровых графиков указывают время ожидания до наступления CR в каждом испытании.Эта взаимосвязь между испытаниями между снижением активности ПК в отдельных веках и появлением следовых количеств CR, по-видимому, аналогична той, которая была продемонстрирована ранее для отсроченного кондиционирования (Halverson et al., 2015a).
Рисунок 3.Гистограммы перистимула и растровые графики, показывающие тесную взаимосвязь между началом CR и снижением активности ПК века. Испытания на растровом графике и поведении расположены в последовательном порядке в A и отсортированы по задержке начала CR (красные точки) в B .Тот же самый пример, отсортированный по времени ожидания до начала поведенческих CR, показан для иллюстрации отношения между испытанием и испытанием между активностью ПК и CR. Для гистограмм и растровых графиков серые полосы представляют длительность тонального стимула, более темная серая линия представляет начало стимуляции век. Для поведенческих разверток синий цвет представляет длительность звукового стимула, а красный – интервал следа. C – E , Поведенческие CRs и профили скорости активации ПК век, отсортированные по группам в соответствии с началом CR и усредненные в пределах каждой ISI.CR (вверху) и данные ПК век (внизу), отсортированные по началу CR (зеленый = самое раннее начало, красный = среднее начало, голубой = самое позднее начало, синий = не-CR). Темно-серые полосы представляют длительность CS, а светло-серые – интервал трассировки. F , G , средняя CR века ( F ) и нормализованная мгновенная скорость стрельбы для всех ПК века ( G ) наложены и выровнены с началом CR (время = 0, пунктир line), записанные во время Delay 500 (оранжевый), Delay 1000 (голубой) и Trace 500/500 (коричневый). H , Те же данные показаны в G , представленные как средняя скорострельность. ПК век снижают активность на ту же величину относительно начала CR во время задержки и кондиционирования следа.
В качестве одного из подходов к преодолению этого артефакта усреднения и количественной оценке взаимосвязи между ПК-ответами век и CR век, мы сгруппировали испытания для каждой обучающей парадигмы в отдельные группы в зависимости от времени начала CR. Диапазоны начала CR были выбраны для создания трех групп (две группы из Delay 1000 из-за меньшего количества зарегистрированных ПК век) с примерно равным количеством испытаний.Поскольку CR с более поздним временем начала, скорее всего, будут иметь меньшую амплитуду, мы добавили дополнительное условие к процедуре группирования, которое гарантировало, что средние значения амплитуд CR были в пределах 10% разницы между группами. Четвертая группа, содержащая испытания без ОК, представляла меньший набор данных, поскольку обычно в этих сессиях было мало испытаний без ОК. На рис. 3, C – E , отдельно для трех парадигм обучения показаны сгруппированные поведенческие и нейронные данные, наложенные друг на друга. Для парадигм кондиционирования Delay 500, Delay 1000 и Trace 500/500 ПК-активность век надежно отслеживала различия между началом CR.Неперекрытие 95% доверительных интервалов средней частоты активации ПК для каждой группы указывает на существенные различия между временем снижения активности ПК, контролирующей CR, с разными средними латентностями начала для всех трех парадигм кондиционирования. Во всех случаях не было изменений в PC-активности век для групп без CR (синие кривые). Существенные различия в ответах ПК века между CR в разное время согласуются с гипотезой о том, что активность ПК века в значительной степени определяет наличие и время CR век как для задержки, так и для следового кондиционирования.
Согласование ПК века и поведенческих реакций на начало ПР обеспечивает более прямую оценку того, как активность ПК века связана с временем ПР. На рис. 3, G и H показаны наложенные средние (рис. 3 G , нормализованные к исходному уровню; рис. 3 H , средняя частота стрельбы) ПК-ответы век для всех трех парадигм обучения. В каждом случае активность ПК века снижается до того же уровня относительно исходного уровня до начала CR (рис. 3 G , H ).При выравнивании по времени начала, профили CR для Delay 500 и Trace 500/500 значительно перекрываются (рис. 3 F ). Точно так же средняя активность ПК века для этих двух форм тренировки также почти неразличима (Рис. 3 G ). В обоих случаях наблюдается небольшое увеличение активности, за которым следует снижение, предшествующее началу CR. Задержка 1000 CR имеет более медленное время нарастания после начала CR и, аналогичным образом, наблюдается более медленное снижение активности ПК века перед началом. Повышение активности перед CR также отсутствует, что, возможно, отражает то, что это увеличение связано с CR с более быстрым временем нарастания.Несмотря на тонкие различия между Delay 1000 и двумя другими парадигмами, одна особенность, общая для всех трех типов ответов, заключается в том, что активность ПК падает до ~ 80% от уровня до CR во время начала CR. Действительно, совпадение 95% доверительных интервалов для всех трех типов ответа согласуется с отсутствием значительных различий между группами (рис. 3 G , H ). Приведенные выше данные демонстрируют сильное сходство между задержкой и условным рефлексом во взаимосвязи между средней ПК-активностью века и временем CR.Изменения в активности ПК век также напрямую связаны с амплитудой CR во время задержки кондиционирования (Halverson et al., 2015a), что мы исследуем далее.
Контроль амплитуды CR по активности ПК века
Чтобы исследовать взаимосвязь между ПК века и амплитудой CR, мы использовали тот же метод группировки, что и для начала CR. На рис. 4, A – C показаны средние профили CR век, сгруппированные по амплитуде CR, с соответствующей средней активностью ПК века для каждой тренировочной парадигмы.Здесь анализ также подтвердил, что средние значения времени начала CR были в пределах 10% разницы между группами. Подобно анализу начала CR, сгруппированная активность ПК в каждой из различных парадигм обучения показала отсутствие перекрытия в 95% доверительных интервалах, что указывает на значительные различия в средней активности ПК, относящейся к CR с разными амплитудами. В дополнение к усредненному сгруппированному анализу, мы также оценили отношения между испытаниями между ПК-активностью века и CR, чтобы выявить возможные различия между задержкой и условным рефлексом, которые может скрыть анализ усредненной активности.Мы сосредоточились на ПК век, записанных во время экспрессии CR во время сеансов Delay 1000 и Trace 500/500, чтобы уравнять ISI по группам. Отношение между испытаниями между амплитудой CR и активностью ПК века для двух примеров показано на рисунке 4 D (черный = задержка 1000, красный = след 500/500). Как для Delay 1000, так и для Trace 500/500, количество всплесков для этих ПК показало сильную линейную связь с амплитудой CR ( r = -0,65 для Delay 1000 и r = -0,63 для Trace 500/500).Был проведен анализ количества пиков в течение последних 500 мс ISI (нормализованного к количеству пиков за 1000 мс до начала CS) в зависимости от амплитуды CR в этом испытании. Точки, сгруппированные около оси y , представляют собой испытания без CR с приблизительно нулевой амплитудой. Рисунок 4 E показывает сводку для каждой записи ПК века для Delay 1000 (черные точки) и Trace 500/500 (красные точки). Связь между активностью ПК века и амплитудой CR была неразличима между Delay 1000 и Trace 500/500.Средние коэффициенты корреляции Пирсона между количеством спайков ПК на веке и амплитудой CR были одинаковыми для Delay 1000 ( r = -0,63 ± 0,09) и Trace 500/500 ( r = -0,62 ± 0,04) и не различались достоверно между парадигмами ( два образца бутстрап-теста, p = 0,61, 10000 повторений) (рис. 4 F ). Средние значения наклонов линейной регрессии также были одинаковыми для разных парадигм (-0,1 ± 0,02 для Delay 1000, -0,081 ± 0,007 для Trace 500/500) и незначительно различались (тест бутстрапа с двумя выборками, p = 0.34, 10000 повторов) (рис.4 G ). Эти данные показывают, что, по крайней мере, с точки зрения амплитуды CR, сходство между ПК века, реагирующим задержкой, и условием трассировки не является результатом усреднения артефактов.
Рис. 4.A – C , Поведенческие профили CR и ПК век, отсортированные по группам в соответствии с амплитудой CR и усредненные в пределах каждого ISI. CR (вверху) и данные ПК век (внизу), отсортированные по максимальной амплитуде CR (голубой = наибольшая амплитуда, красный = средняя амплитуда, зеленый = наименьшая амплитуда, синий = не-CR).Темно-серые полосы представляют длительность CS, а светло-серые – интервал трассировки. D , Корреляция между испытаниями между амплитудой CR и количеством спайков, нормализованными к базовой линии от ПК век, записанных во время Delay 1000 и Trace 500/500. Каждая точка представляет собой нормализованное количество всплесков за последние 500 мс перед США как функцию амплитуды CR в отдельных испытаниях. Точки и линия наилучшего линейного соответствия соответствуют Delay 1000 (черный) и Trace 500/500 (красный) индивидуальному веку PC. Пример s . E , Сводка корреляций между испытаниями между нормализованным количеством спайков и амплитудой CR для всех ПК век, записанных во время Delay 1000 и Trace 500/500. F , Распределение коэффициентов корреляции между количеством спайков и амплитудой CR. Каждая точка представляет ПК с одним веком во время Delay 1000 и Trace 500/500. Планки погрешностей указывают на SEM. G , аналогично F , но сравнивая наклоны линейной регрессии из анализов в E .
Отслеживание одного и того же нейрона от задержки до кондиционирования трассы
В двух случаях мы смогли проследить один и тот же ПК века от кондиционирования задержки через более поздние сеансы согласования трассы, где реакция трассировки была на асимптоте. Критерии идентификации ПК как одного и того же во всех сеансах не зависели от его ответов, связанных с испытаниями, и вместо этого основывались на согласованности относительного расположения кластера между сеансами, формы сигнала по четырем каналам тетрода и гистограмм ISI.Записи с одного и того же тетрода, вероятно, были с одного и того же ПК в течение сеансов в тех случаях, когда каждая из этих переменных была согласованной.
Один из примеров включал ПК века, за которым наблюдали от выражения условного рефлекса задержки (Delay 500 хорошо натренирован; рис. 5, внизу) до хорошо рассчитанного по времени экспрессии CR Trace 500/500 (рис. 5, вверху). Гистограммы перистимула, растровые графики и поведение показаны для пяти сеансов, когда кролик переходил от задержки к кондиционированию трассировки. Наличие или отсутствие CR и их время всегда сопровождалось параллельными изменениями в ответах ПК века.Устойчивый (хорошо обученный) отклик с задержкой сопровождался значительным снижением активности ПК. В сеансе перехода от Delay 500 к Trace 500/500 исчезновение CR задержки происходило параллельно с исчезновением снижения активности ПК. Неизвестно, вызваны ли CR с ранней задержкой в следующем сеансе после сеанса перехода спонтанным восстановлением погашенных откликов задержки или началом ответа трассировки; так или иначе, ответы сопровождались снижением активности ПК в соответствующее время.Наконец, в последнем сеансе (хорошо обученные трассы 500/500) устойчивые CR трассировки показали более поздние начала по сравнению с более ранним временем начала, наблюдаемым во время кондиционирования задержки. Более позднее время начала CR во время кондиционирования следов сопровождалось соответственно более поздним снижением активности ПК. Чтобы более четко показать взаимосвязь между снижением активности ПК века и CR во время кондиционирования следа, данные хорошо тренированного сеанса также показаны с сортировкой по началу CR (рис. 5, вверху). Этот пример показывает, что снижение активности ПК одного века сдвигается, чтобы соответствовать изменениям во времени условных реакций века, выраженных во время задержки и кондиционирования следа.
Рисунок 5.Записи с ПК с одним веком, проведенные в течение 5 дней, необходимых для перехода от экспрессии хорошо обученных CR во время задержки 500 (внизу) к Trace 500/500 (вверху). Показаны гистограммы перистимула (интервалы 10 мс) и растровые графики в порядке каждого 108 пробных сеансов с соответствующими ответами век. Для поведенческих разверток синий представляет длительность тонального сигнала CS 500 мс, а красный – интервал трассировки 500 мс. Данные хорошо обученного сеанса Trace 500/500 показаны в порядке сеанса и снова отсортированы по задержке CR (красные точки) (вверху), чтобы более четко показать взаимосвязь между снижением активности ПК века и CR.
Во втором случае мы смогли проследить ПК с одним веком от выражения CRs Delay 1000 с CS тона 9,5 кГц посредством сбора данных и надежного выражения CR во время Trace 500/500 с CS тоном 1 кГц. Рис. 6 A показывает гистограммы перистимула, растровые графики и ответы век из четырех из восьми сеансов записи, в которых отслеживался этот компьютер век. Четыре сеанса, показанные на Рисунке 6 A , снизу вверх: первый сеанс Delay 1000, четвертый сеанс Delay 1000, сеанс перехода от Delay 1000 к Trace 500/500 с другой CS и четвертый сеанс Прослеживание 500/500 кондиционирования.Чтобы наглядно проиллюстрировать, что взаимосвязь между ПК-активностью века и условными ответами века не меняется для трассировки по сравнению с откликами задержки, мы построили комбинированный растровый график ПК-данных из рисунка 6 A , отсортированных по началу CR с началом Trace 500 / 500 ответов обозначены красными точками, а начало задержки 1000 ответов обозначено синими точками. Как для задержки, так и для отслеживания ответов, снижение активности ПК век происходит непосредственно перед началом ОК, и испытания без ОК для каждой обучающей парадигмы не сопровождаются изменениями активности ПК (нижняя часть растра).Этот компьютер продемонстрировал аналогичную взаимосвязь с поведенческими реакциями в обеих парадигмах и показывает, что один компьютер демонстрирует снижение, связанное с обучением, для разных CS.
Рисунок 6.A , Пример ответов от ПК с одним веком, которые были записаны на протяжении перехода от выражения хорошо обученных CR с задержкой 1000 к экспрессии CR с помощью Trace 500/500 (и другого тона CS). Гистограммы перистимула (интервалы 10 мс), растровые графики и реакции век показаны в хронологическом порядке (снизу вверх) для двух сеансов Delay 1000 и двух Trace 500/500.Поведенческие развертки показывают, как время CR изменялось во время перехода от задержки к условию трассировки. Синий представляет длительность тонального сигнала CS, а красный – продолжительность интервала трассировки. Обратите внимание на тесное соответствие между испытаниями между снижением активности ПК век и началом полного ответа на каждом сеансе. B , Все испытания из сеансов, показанных в A , были объединены в единый растровый график и отсортированы по началу CR. Это было сделано, чтобы еще раз продемонстрировать постоянную взаимосвязь между ответами ПК века и CR века как во время задержки, так и во время отслеживания кондиционирования века.Испытания с началом CR, обозначенные синими точками, представляют испытания с задержкой 1000, а красные точки представляют испытания с отслеживанием 500/500. Черная линия в момент времени = 0 представляет начало CS.
CR, полученные в результате кондиционирования трассировки, лучше рассчитаны по времени (показывают более поздние задержки до начала и более быстрое время нарастания), чем отклики с задержкой с тем же ISI (Kalmbach et al., 2010a). Данные свидетельствуют о том, что это опосредовано способностью мозжечка научиться реагировать на постоянные входные сигналы и подавлять реакцию на входные сигналы CS во время кондиционирования следа, что помогает отсрочить наступление CR после смещения CS (Kalmbach et al., 2009, 2010b). Это улучшение во времени очевидно в примере перехода от кондиционирования с задержкой 1000 к кондиционированию с отслеживанием 500/500, показанного на фиг. 6, , , как в поведенческих CR, так и в ответах ПК века. Сортировка этих данных по латентности до начала CR показывает улучшенное время (CR и ответы ПК век) от сеанса перехода от Delay 1000 до Trace 500/500 кондиционирования (рис. 7 B ) к хорошо обученному сеансу кондиционирования трасс (рис. . 7 А ).Чтобы количественно оценить это изменение во времени CR, мы рассчитали отдельные совокупные распределения вероятностей времени начала CR для двух сеансов задержки по сравнению с двумя сеансами трассировки из данных, показанных на рисунке 6 (рис. 7 C ). Двухвыборочный тест Колмогорова – Смирнова показал, что эти распределения существенно различались ( p <0,0001, α = 0,01). Эти результаты демонстрируют, что эта поведенческая парадигма обеспечивает подлинный переход от отклика Delay 1000 с его характерной более низкой синхронизацией к реагированию Trace 500/500, где CR более точно рассчитываются по времени при одном и том же общем ISI.Отсортированные растровые графики на рисунке 7, A и B , качественно предполагают, что, несмотря на изменение времени CR, взаимосвязь между ответами ПК века и CR не изменяется, что согласуется с гипотезой о том, что активность ПК века контролирует экспрессию CR. аналогично для трассировки и для кондиционирования задержки.
Рисунок 7.A , B , гистограммы перистимула (интервалы 10 мс), растровые графики и CR век, отсортированные по началу CR (красные точки), показаны для перехода ( B ) и хорошо обученный ( A ) Проследите 500/500 сеансов с рисунка 6.Эти два сеанса иллюстрируют улучшение задержки (времени) начала CR во время переключения с Delay 1000 на Trace 500/500. Обратите внимание на количество CR, которые начинаются во время CS в переходном сеансе кондиционирования трассировки, тогда как очень немногие CR начинаются во время CS во время хорошо обученного сеанса. C , Кумулятивное распределение вероятности задержки начала CR для сеансов Delay 1000 (черный) и Trace 500/500 (красный) из рисунка 6, иллюстрирующего более позднюю задержку начала во время кондиционирования Trace 500/500 по сравнению с задержкой 1000 при том же общий ISI.
Предсказания обратной модели активности ПК во время задержки и кондиционирования трассы
Выше мы продемонстрировали, что взаимосвязь между связанными с обучением уменьшениями ПК век и началом CR, а также корреляция между амплитудой CR и количеством спайков ПК согласованы для обеих задержек. и следовое кондиционирование. Одним из наиболее убедительных доказательств того, что связанные с обучением изменения ПК контролируют выражение условных ответов век (при задержке кондиционирования), является тесная взаимосвязь между активностью ПК века и кинематическими свойствами CR.Связь или ее отсутствие между активностью ПК века и реакциями следов представляет собой серьезный тест того, насколько похожи или различны вклады коры мозжечка в задержку по сравнению со следовыми CR. Это ставит под вопрос, согласуются ли отношения между временным профилем скорости стрельбы ПК век и кинематикой CR как функцией времени в парадигмах задержки и следа, которые имеют центральное значение. Мы проверили этот вопрос, используя подход обратной модели, который ранее использовался для моделирования взаимосвязи между активностью ПК и поведением в исследованиях плавного преследования (Medina and Lisberger, 2007, 2009).В обратной модели профиль скорости стрельбы ПК моделируется как линейная взвешенная сумма положения века, скорости и ускорения как функции времени в дополнение к базовой константе скорости. Веса в модели ( a – c в уравнении) относятся к относительному вкладу положения, скорости и ускорения во временной профиль скорости стрельбы.
Данные, использованные для подбора модели, и данные, использованные для прогнозирования скорости стрельбы ПК из модели, были основаны на отдельных сериях испытаний (см. Материалы и методы).Точность модели определялась величиной разброса фактической средней скорости срабатывания ПК для века в тестовых испытаниях, которая была зафиксирована прогнозом. На рисунке 8 показана обратная модель, примененная к записи ПК каждого века для Delay 1000 ( n = 8) и Trace 500/500 ( n = 26). Примеры записей ПК отдельных век во время задержки 1000 (рис. 8 A ) и трассировки 500/500 (рис. 8 B ) показывают результаты моделирования для двух примеров отдельных единиц.Средняя фактическая скорость срабатывания ПК века (коричневые линии) из различных образцов тестовых испытаний из сеанса, а также прогнозируемая скорость срабатывания ПК модели (оранжевые линии) показаны вверху. Величина отклонения фактической скорости срабатывания ПК, зафиксированная прогнозом, показана для каждого ПК, записанного во время задержки 1000 и трассировки 500/500 на рисунке 8 C . Обратная модель обеспечивает точные прогнозы ответов ПК века для Delay 1000 ( R 2 = 0,66 ± 0.08) и Trace 500/500 ( R 2 = 0,60 ± 0,04). Поскольку количество тестовых испытаний, используемых для генерации каждого прогноза, было небольшим (среднее значение = 20 на итерацию), дисперсия, не зафиксированная моделью, вероятно, происходит из-за случайных колебаний скорости стрельбы вокруг средней базовой скорости от испытания к испытанию. Точность прогноза существенно не отличалась для популяций ПК век от парадигм задержки и трассировки (тест начальной загрузки с двумя выборками, p = 0,23, 10 000 повторений).
Рисунок 8.Анализ обратной модели ПК-активности век во время Delay 1000 и Trace 500/500. A , B , Примеры из отдельных ПК век модели обратной скорости стрельбы для Delay 1000 ( A ) и Trace 500/500 ( B ). Профиль средней скорости стрельбы ПК (коричневые линии) на четырех итерациях тестовых испытаний (см. Материалы и методы), а также прогнозируемая скорость стрельбы из обратной модели (оранжевые линии). Тонкие линии представляют результаты четырех итераций тестовых испытаний, а толстые линии представляют собой среднее значение по ним.Вклады положения века, скорости и ускорения в прогнозируемую скорость стрельбы ПК показаны внизу. Темно-серая заштрихованная область представляет длительность тонального стимула, а светло-серая область представляет собой интервал следа. C , Величина отклонения временного профиля фактической скорострельности ПК, зафиксированная обратной моделью. Каждая точка представляет собой цвет ПК одного века, кодированный в соответствии с парадигмой обучения. Гистограмма показывает среднее значение R 2 для всех записей ПК в рамках парадигмы.Планки погрешностей указывают на SEM. D – F , Взаимосвязь между положением, скоростью и весами ускорения в обратной модели. Каждая точка представляет собой цвет ПК одного века, кодированный в соответствии с парадигмой обучения. G – I , Кумулятивные распределения вероятностей весов каждой кинематической переменной в обратной модели, построенные отдельно для каждой парадигмы обучения.
Прогноз скорости срабатывания ПК для века с помощью обратной модели основан на линейной комбинации профилей положения века, скорости и времени ускорения.Вклады в положение века, скорость и ускорение показаны в нижней части рисунка 8, A и B для Delay 1000 и Trace 500/500, соответственно. В каждом случае тонкие линии соответствуют различным образцам тестовых испытаний, а более толстая линия представляет среднее значение по каждому из образцов. Для обоих примеров ПК из сеансов задержки и трассировки положение век и скорость обеспечивают аналогичный вклад в прогнозируемую скорость стрельбы, тогда как компонент ускорения дает незначительный вклад.Как соответствующие веса распределяются по популяциям ПК век для Delay 1000 и Trace 500/500, показано на Рисунке 8, D – F . Веса положения, скорости и ускорения были нормализованы так, чтобы сумма их абсолютных значений была равна единице. Эта нормализация позволила изучить возможные различия между тем, как ПК век кодируют кинематику задержки и отслеживают CR с одним и тем же интервалом. Диаграммы разброса (рис. 8 D – F ) показывают, что как во время задержки, так и во время кондиционирования трассировки, положение век и вклад скорости являются доминирующими, тогда как веса ускорения век являются номинальными (рис.8 D – F , все точки разбросаны по оси x для парадигм задержки и трассировки). Кумулятивные распределения вероятностей каждой кинематической переменной веса по всем зарегистрированным ПК век во время парадигм Delay 1000 и Trace 500/500 показаны на Рисунке 8, G – I . Хотя существуют вариации в зависимости от положения и скорости вкладов ПК отдельных век, не было значительных различий в весах кумулятивной функции распределения между задержкой и условием следа (двухвыборочный тест Колмогорова-Смирнова, p = 0.82, p = 0,85 и p = 0,94 для рис. 8, G – I соответственно). Эти результаты показывают, что не только ПК век управляют кинематикой задержки и отслеживания CR, рассчитанных на один и тот же интервал (ISI 1000), но и то, как эта кинематика связана с активностью ПК, неразличима в двух парадигмах обучения.
Обратная модель предсказывает активность ПК через кондиционирование задержки и следа. следы кондиционирования век.Если взаимосвязь между ПК-активностью века и CR одинакова между задержкой и кондиционированием следа, тогда должна быть возможность получить веса, полученные из тестовых испытаний во время кондиционирования задержки, чтобы затем предсказать активность ПК века во время кондиционирования следа (и использовать веса следа для прогнозирования задержки ) с той же моделью скорострельности. Мы реализовали этот тест для ПК века, показанного на рисунке 6. Мы полагаем, что эти данные взяты с того же ПК века, проведенного в течение многих сеансов, но это предположение не является необходимым для того, чтобы сделать выводы ниже.
Следуя этой логике, мы сначала использовали часть испытаний из одной парадигмы (например, Trace 500/500), чтобы подогнать модель, а затем применили результаты подбора к тестовым испытаниям в рамках той же парадигмы (рис. 9 ). A , слева) или через парадигму до Delay 1000 (рис.9 A , справа). Коричневые линии на рис. 9 A соответствуют фактической средней скорости срабатывания ПК во время тестовых испытаний, тонкие линии соответствуют каждой итерации выбранных тестовых испытаний, а более толстая линия представляет среднее значение.Оранжевые линии – это предсказания обратной модели на основе весов, полученных из данных соответствия для положения, скорости и ускорения, плюс константа базовой линии. На рис. 9 B показана та же процедура с использованием обратной модели для прогнозирования активности ПК во время кондиционирования Trace 500/500 на основе весов, полученных в сеансах кондиционирования Delay 1000. Межпарадигмальные прогнозы скорости стрельбы ПК не только воспроизводили общую динамику выученного снижения активности, но также фиксировали большую часть фактической дисперсии скорости стрельбы ( R 2 = 0.56 ± 0,05 для задержки от трассы, Рис. 9 C , черная полоса; R 2 = 0,63 ± 0,05 для кривой от задержки, рис.9 D , красная полоса). Способность модели воспроизводить большую часть данных ПК в разных парадигмах особенно убедительна, учитывая тонкие кинематические различия, наблюдаемые во время выражения CR Delay 1000 и Trace 500/500. При прогнозировании разных парадигм наблюдалась значительная разница в величине дисперсии, зафиксированной в модели (двухвыборочный бутстрап-тест, p <0.001, черные и красные полосы на рис.9 ( C , D ). Разница между парадигмами, вероятно, была вызвана двумя факторами. Во-первых, как подробно описано выше, основное различие между Delay 1000 и Trace 500/500 связано с кинематикой CR, которая использовалась для создания прогнозов с использованием подхода обратной модели. Во-вторых, модель смогла учесть большое количество вариаций внутрипарадигмальных прогнозов в обоих случаях. Следовательно, межпарадигмальное различие в точности прогнозов, вероятно, является отражением высокой точности внутрипарадигмальных прогнозов в сочетании с тонкими кинематическими различиями между двумя парадигмами, а не взаимосвязью активности ПК с CR.Эти результаты предоставляют дополнительные доказательства в поддержку общих результатов, указывающих на то, что взаимосвязь между скоростью возбуждения ПК века и кинематикой CR одинакова для задержки и кондиционирования следов.
Рис. 9.Обратная модель использовалась для прогнозирования активности ПК века как внутри, так и между парадигмами. Все данные, используемые для этого анализа, взяты из сеансов, показанных на рисунке 6. A , активность ПК и кинематика век, собранные во время Trace 500/500, были использованы для подгонки обратной модели.Результаты каждой подгонки применялись для прогнозирования срабатывания ПК во время тестовых испытаний для сессий Trace 500/500 (в рамках парадигмы) и Delay 1000 (в рамках всей парадигмы). Цветовая кодировка такая же, как на рисунке 8. Обе парадигмы нанесены на график, показывающий фактическую (коричневые линии) и прогнозируемую (оранжевые линии) частоту срабатывания ПК, усредненную по результатам испытаний. B , То же, что и A , но данные из сеансов Delay 1000 использовались для подгонки обратной модели, а затем для прогнозирования как внутри парадигмы, так и между парадигмами. C , D , величина отклонения фактического временного профиля скорости воспламенения ПК века, зафиксированная обратной моделью. В каждом случае первая полоса представляет точность прогнозирования в рамках парадигмы (например, трасса прогнозирования трассировки), а вторая полоса показывает результаты прогнозирования в рамках парадигмы (например, задержку прогнозирования трассы). Здесь каждая точка представляет значение R, 2 для каждой итерации пробного испытания, а гистограмма показывает R 2 , усредненные по итерациям.Планки погрешностей указывают на SEM.
Обсуждение
Мы проверили гипотезу, выдвинутую на основе определенных исследований поражений и нокаутов, что кора мозжечка либо не участвует в формировании следового состояния век, либо, по крайней мере, что вовлечение коры мозжечка принципиально различается между задержкой и следом. кондиционирование век (Woodruff-Pak et al., 1985; Kishimoto et al., 2001a, b, c; Takehara-Nishiuchi, 2018). Чтобы проверить эту гипотезу, мы записали ответы ПК из области коры мозжечка, которые, как известно, необходимы для приобретения и своевременной экспрессии CR во время задержки и следового кондиционирования (Perrett et al., 1993; Гарсия и др., 1999; Kalmbach et al., 2010a; Halverson et al., 2015a). Мы обнаружили, что ответы ПК век как на задержку, так и на следы CR соответствуют гипотезе о том, что одни и те же ПК или, по крайней мере, ПК в одной и той же небольшой области коры мозжечка контролируют экспрессию CR во время как задержки, так и следового кондиционирования. Связь между изменениями активности ПК век и поведенческим выражением CR век почти неразличима в двух парадигмах. Эти данные не согласуются с гипотезами о том, что область коры мозжечка, связанная с экспрессией CRs задержки, не играет той же роли во время кондиционирования следовых век и что кора мозжечка необходима для задержки, но не для кондиционирования следов.Напротив, эти данные подтверждают гипотезу о том, что связанные с обучением изменения в ПК контролируют экспрессию как следовых, так и отсроченных CR век, а правила, действующие в коре мозжечка, включая то, необходимо ли это для обучения и экспрессии CR, не различаются для следов. и отсрочить кондиционирование. В сочетании с предыдущими результатами (Kalmbach et al., 2009, 2010b), настоящие результаты предполагают, что основное различие для мозжечка между задержкой и следами кондиционирования век заключается в сложности входов мшистых волокон.При кондиционировании отсрочки со слуховым CS необходимый вход CS управляется проекцией медиального слухового таламуса на мост (Halverson and Freeman 2006, 2010; Halverson et al., 2008, 2010, 2015b), тогда как при кондиционировании следа век происходит дополнительный вход, передающий стойкие ответы на мосты через передний мозг или таламические структуры. Хотя это различие может объяснить лучшее время, наблюдаемое в следовых CR, оно не требует принципиально других мозжечковых процессов или правил обучения.
Одно небольшое различие было очевидным в ответах ПК век для Delay 1000 по сравнению с Delay 500 и Trace 500/500. В Delay 500 и Trace 500/500, где скорости CR несколько выше, наблюдалось небольшое увеличение активности PC сразу после начала CS и перед началом CR (Рис. 2 C ). Это увеличение не наблюдалось в ответах ПК век во время задержки 1000. Предыдущая работа, включающая как эмпирические данные, так и моделирование мозжечка, предполагала, что увеличение активности ПК на ранней стадии CS может улучшить время за счет активного подавления активности в глубоких ядрах мозжечка (Medina et al. ., 2000). Отсутствие начального увеличения ПК-активности века во время задержки 1000 может быть связано с худшим временем, наблюдаемым, когда кролики обучаются более длительным ISI по сравнению с Delay 500 и Trace 500/500 (Kalmbach et al., 2010b).
Первоначальное предположение, что кора мозжечка не является необходимой для кондиционирования следов, возникло из эксперимента, показывающего, что большие аспирационные поражения коры мозжечка не влияли на экспрессию CR во время кондиционирования следов (Woodruff-Pak et al., 1985). Ранее аналогичные выводы были сделаны для отсрочки кондиционирования век на основании исследований, в которых сообщалось, что поражения коры мозжечка не вызывают сдвигов латентности CR или полностью устраняют CR (Yeo et al., 1984, 1985а; Лавонд и Стейнмец 1989; Йео и Хардиманн 1992; Кларк и Лавонд 1994; Attwell et al., 1999). Позже исследования повреждений и инактивации выявили более латеральную и вентральную область коры мозжечка (между HVI и HV передней доли), которая необходима для своевременной экспрессии CR как для задержки, так и для следового кондиционирования, и которая необходима для приобретения и исчезновения CR век для задержки кондиционирования (Perrett et al., 1993; Perrett and Mauk 1995; Garcia and Mauk 1998; Garcia et al., 1999; Kalmbach et al., 2010a). Мозжечка также достаточно для изучения CR век в протоколе отслеживания, когда одно входящее мшистое волокно стимулируется в течение продолжительности CS, а стимуляция отдельного входящего мшистого волокна сохраняется в интервале следа (Kalmbach et al., 2010b). Следовательно, как для отсрочки, так и для следового кондиционирования, эксперименты с ранними повреждениями, которые не смогли выявить влияния на время, приобретение и угасание, могли просто пропустить критическую область коры мозжечка (Nores et al., 2000).
Способность делать выводы о мозжечковых процессах и функциях на основе исследований кондиционирования век, включая необходимость коры мозжечка, основана на предположении, что CR управляются исключительно мозжечковой продукцией. В отношении кроликов это предположение подтверждается множеством сходных данных (McCormick and Thompson 1984; Lavond et al., 1984; Yeo et al., 1985b; Berthier, Moore 1990; Krupa et al., 1993; Krupa and Thompson 1997; Garcia и Mauk 1998; Choi and Moore 2003; Kalmbach et al., 2010a; Halverson et al., 2015a). Статус этого предположения для мыши значительно менее определен. Хотя Heiney et al. (2014) продемонстрировали, что CR на веках мышей зависят от мозжечка при определенных условиях обучения, другие недавние исследования продемонстрировали, что CR на веках у мышей могут быть вызваны нецеребеллярными процессами (Boele et al., 2010; Sakamoto and Endo 2010; Schonewille et al., 2011; Siegel et al., 2012). В той степени, в которой это правда, демонстрация щадящего обучения у мышей с дегенерирующими ПК или с дефицитом LTD в синапсах на ПК (Kishimoto et al., 2001а, б, в; Браун и др., 2010; Yang et al., 2015) может не иметь никакого отношения к необходимости этих процессов в обучении мозжечка. Такие выводы требуют дополнительных мер контроля, таких как демонстрация того, что CR у этих мышей опосредуются мозжечком – что они, например, устраняются повреждениями глубоких ядер мозжечка. Во многих из этих исследований поведенческие атрибуты обучения указывают на высокую вероятность того, что CR опосредуются нецеребеллярными процессами и даже неассоциативными процессами.
Задержка и кондиционирование следа различаются только количеством входов в мозжечок, необходимых для обучения и экспрессии CR. Основываясь на наших компьютерных записях век, США задействуют одну и ту же область века нижней оливы во время как задержки, так и трассировки. Эти входные сигналы восходящего волокна затем диктуют ПК, которые будут участвовать в обучении и выполнении CR для любой задачи (трассировки или задержки), связанной с реакциями век. В дополнение к восходящему входу волокна, для кондиционирования задержки требуется входной сигнал из замшелого волокна от CS, тогда как для кондиционирования трассы требуется дополнительный постоянный вход для преодоления временного разрыва (интервала трассы) между смещением CS и США (Clark and Squire 1998; Siegel et al., 2012). Мозжечок обучается постоянному входу во время кондиционирования следа, в то же время подавляя поведенческие реакции на вход CS (тональный сигнал). Этот механизм обучения приводит к CR во время подготовки трассы, которые показывают лучшее время (т.е. более длительную задержку до начала) по сравнению с CR задержки, обученными тому же общему интервалу (Kalmbach et al., 2011). Мы предполагаем, что у кроликов данные однозначно подтверждают гипотезу о том, что как задержка, так и отслеживание состояния век одинаково зависят от коры мозжечка.Повреждения и обратимая инактивация области век коры мозжечка производят сравнимые эффекты как для задержки, так и для отслеживания кондиционирования век (Kalmbach et al., 2010a), входящие волокна лазания определяют, какая область коры мозжечка контролирует обучение, и настоящие записи показывают, что ПК из эта область одинаково связана с отслеживанием и задержкой обусловленности присутствием, временем и амплитудой условных реакций.
Модель кондиционирования века Павлова, основанная на синаптической организации мозжечка.
- М Д Маук и
- N H Донеган
- Кафедра нейробиологии и анатомии, Медицинская школа Техасского университета в Хьюстоне 77030, США.
Аннотация
Мы представляем модель, основанную на синаптической и клеточной организации мозжечка, для получения разнообразных явлений. наблюдается в павловском кондиционировании век.Эти явления рассматриваются с точки зрения критических путей и свойств сети, а также сайты и правила синаптической пластичности. Теория основана на четырех основных гипотезах: (1) Два мозжечка. участки пластичности участвуют в формировании кондиционирования: (а) двунаправленная долговременная депрессия / потенциация в синапсах гранулярных клеток на клетки Пуркинье (gr -> Pkj) в коре мозжечка и (б) двунаправленная пластичность в межположительном ядре, которое контролируется ингибирующими входами от клеток Пуркинье; (2) активность лазящих волокон регулируется до равновесного уровня при что чистая сила синапсов gr -> Pkj остается постоянной, если не представлен неожиданный безусловный стимул (US) или ожидаемый US опущен; (3) изменяющееся во времени представление условного раздражителя (CS) в коре мозжечка.